- ホーム
- >
- 細胞生物学用語集トップ
- >
- 新着細胞生物学用語集(%EF%BF%BD%EF%BF%BD%EF%BF%BD%EF%BF%BD%EF%BF%BD%EF%BF%BD%EF%BF%BD%EF%BF%BD%EF%BF%BD%EF%BF%BD%EF%BF%BD%EF%BF%BD%EF%BF%BD%EF%BF%BD%EF%BF%BD%EF%BF%BD%EF%BF%BD%EF%BF%BD%EF%BF%BD%EF%BF%BD%EF%BF%BD%EF%BF%BD%EF%BF%BD%EF%BF%BD%EF%BF%BD%EF%BF%BD%EF%BF%BD%EF%BF%BD%EF%BF%BD%EF%BF%BD%EF%BF%BD%EF%BF%BD%EF%BF%BD)
新着細胞生物学用語集(%EF%BF%BD%EF%BF%BD%EF%BF%BD%EF%BF%BD%EF%BF%BD%EF%BF%BD%EF%BF%BD%EF%BF%BD%EF%BF%BD%EF%BF%BD%EF%BF%BD%EF%BF%BD%EF%BF%BD%EF%BF%BD%EF%BF%BD%EF%BF%BD%EF%BF%BD%EF%BF%BD%EF%BF%BD%EF%BF%BD%EF%BF%BD%EF%BF%BD%EF%BF%BD%EF%BF%BD%EF%BF%BD%EF%BF%BD%EF%BF%BD%EF%BF%BD%EF%BF%BD%EF%BF%BD%EF%BF%BD%EF%BF%BD%EF%BF%BD)
| 非対称分裂 |
|---|
| 【Asymmetric cell division】 |
| 入江 賢児 |
| 筑波大学大学院人間総合科学研究科(基礎医学系)分子細胞生物学グループ |
| お問合せ |
細胞分裂によって2つの異なる細胞を生じるような細胞分裂。非対称分裂は細胞の多様性を生みだす基本的なしくみである。個体発生の過程では、少数の未分化な前駆細胞(もしくは幹細胞)が細胞増殖を繰り返し、特定の機能や構造を形成する細胞集団が形成されていく。非対称分裂は、このような過程で、細胞内の非対称性や細胞外の微細環境(ニッチ、niche)に基づいて、未分化な細胞からより分化した細胞を生じるメカニズムである。
参考文献1;Mechanisms of asymmetric cell division: two Bs or not two Bs, that is the question. Horvitz HR, Herskowitz I. Cell. 1992 Jan 24;68(2):237-55.
|
| 酵母の紡錘体微小管 |
|---|
| 【Spindle microtubules in yeast】 |
| 佐藤 政充 |
| 東京大学大学院理学系研究科生物化学専攻 |
| お問合せ |
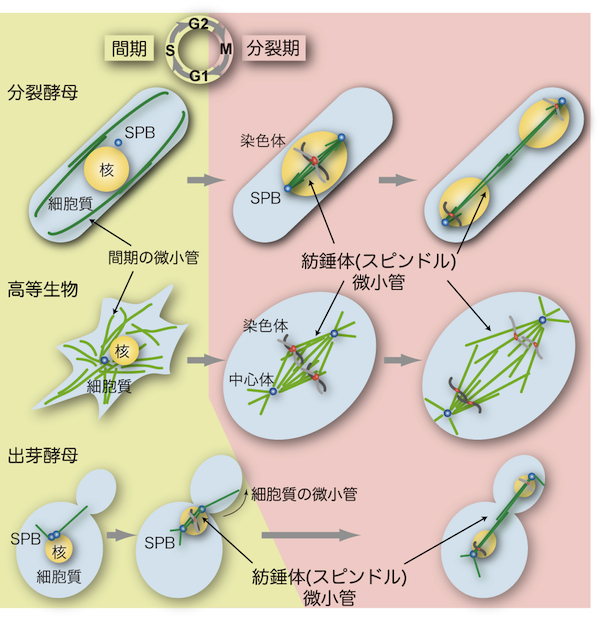
| 紡錘体(スピンドル、spindle)は微小管(microtubule)の束が多数集まって構成される細胞内の構造である。真核細胞において紡錘体はゲノム染色体を2個の娘細胞に分配するために必要不可欠の役割を担う(「紡錘体微小管」の項を参照)。酵母における紡錘体微小管の基本的な機能や性質は高等生物のものと良く似ているが、相違点もいくつかある。 高等生物では一般的に紡錘体はひし形であるが、酵母では微小管が束ねられて直線状の形を示す(図参照)。また、酵母では分裂期でも核膜が崩壊しない(closed mitosisと呼ばれる)ため、紡錘体が核内に形成されるという大きな特徴がある(「酵母の核分裂」の項を参照)。 分裂酵母では、間期(interphase)に見られる細胞質の微小管構造は分裂期(mitosis)になると消失し、かわりに核内に紡錘体微小管を形成する(図参照)[1]。このように、細胞周期の時期に応じて、微小管の形成される場所が細胞質から核へと移る。その分子メカニズムはまだ明らかにされていないが、第一に微小管を安定化するタンパク質が核内に蓄積することが重要であり、第二に微小管を形成する原点であるスピンドル極体(SPB;「酵母の中心体」の項を参照)が分裂期に核膜に埋め込まれることが必要であると考えられる[2, 3]。 これに対して、出芽酵母のSPBは細胞周期を通して常に核膜に埋め込まれているため(「酵母の中心体」の項を参照)、間期でも核内にわずかながら微小管構造が存在し[4]、分裂期に突入する前のG2期の段階で紡錘体を形成する(図参照)。さらに出芽酵母では、出芽した娘細胞に核を正しく分配させるために、細胞質の微小管が細胞の内壁をたどっていき紡錘体を回転させ(spindle orientation、図参照)、核を娘細胞へと誘導する。 紡錘体微小管は、複製された染色体の動原体部位(キネトコア、kinetochore)を接着して両極に引っ張ることで染色体を分配する。微小管の接着が未完了の場合や、不適切な接着が起きた場合には、紡錘体形成チェックポイント(spindle assembly checkpoint)と呼ばれる監視機構が活性化して、すべての動原体が微小管によって正しく捕捉されるまで細胞周期を分裂中期(metaphase)に停止させる。Mad2やBub1などのチェックポイント因子は酵母からヒトまで真核細胞生物で幅広く保存されている。 参考文献 |
| 酵母の核分裂 |
|---|
| 【Closed mitosis: nuclear division in yeast】 |
| 新井 邦生・佐藤 政充 |
| 東京大学大学院理学系研究科生物化学専攻 |
| お問合せ |
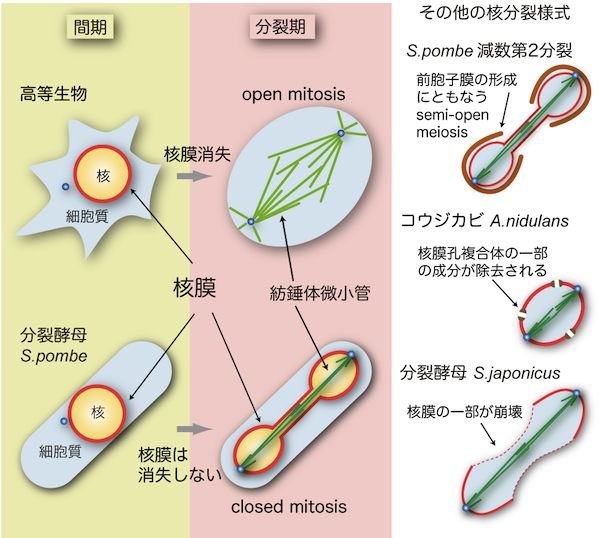
| 酵母における細胞分裂は、基本的には高等生物の細胞分裂と類似しているが、分裂期に核膜が崩壊しないことが構造上の最も大きな違いである。
高等生物では、分裂期に核膜が消失する(nuclear envelope break down, NEBD)ため、その形態はopen mitosisと呼ばれる。酵母を含む多くの単細胞生物では分裂期でも核膜が崩壊せず、closed mitosisと呼ばれる(図参照)[1]。したがって、Ran GTPaseによって統括される核・細胞質間の物質輸送は分裂期にも機能する。紡錘体微小管は核内に形成される必要がある(「酵母の紡錘体微小管」「酵母の中心体」の項を参照)が、Ranが輸送する重要な積み荷として、紡錘体微小管の形成に重要な役割を果たすAlp7/TACC (transforming acidic coiled-coil)タンパク質が知られている[2]。分裂後期(anaphase)には、紡錘体微小管が伸長することで核膜が維持されたまま核が2個に分裂する。このように、体細胞分裂において核分裂をおこすためには紡錘体微小管の伸長が不可欠である。 酵母の核分裂はclosed mitosisのみだと長らく信じられてきた。しかし近年、分裂酵母S. pombeの減数第2分裂においては、核膜は崩壊しないものの、核・細胞質間を隔てる機能が失われることが分かった(semi-open meiosis, virtual open meiosisと呼ぶ;図参照)[3,4]。減数第2分裂では配偶子(胞子)を作るために、核膜のまわりに前胞子膜(forespore membrane)と呼ばれる膜を形成する。前胞子膜が形成されることで、核膜あるいは核膜孔複合体(nuclear pore complex)にも連動して影響が生じている可能性がある。 コウジカビA. nidulansは分裂期に核膜孔複合体の一部の成分が除去されることで核・細胞質間の隔たりが無効化されるsemi-open mitosisをおこなう[5]。また、S. pombe近縁種の分裂酵母S. japonicusでは、体細胞分裂時に核膜の一部が崩壊することが分かった(図参照)[6]。このように、酵母やカビに限っても核分裂には様々な様式があることが明らかになってきた。それぞれの様式にどのような利点があるのかは推測の域を出ないが、各生物の生態に密接に関連した様式の核分裂をおこなっているのかもしれない。 参考文献 |
| 酵母の中心体:スピンドル極体 |
|---|
| 【SPB: spindle pole body】 |
| 戸谷 美夏 |
| 理化学研究所 神戸研究所 発生・再生科学総合研究センター (CDB) |
| お問合せ |
| 酵母は、動物細胞における中心体に相当する細胞内小器官として、スピンドル極体 (SPBと呼ばれることが多い)をもつ。SPBは、直径約100nm程度の円盤が重なったような、中心体とは大きく異なる構造をもつが、中心体と同様に、細胞内で微小管形成中心(MTOC: microtubule organizing center)として働いている(図参照)。SPBと中心体には、微小管の形成起点となるγチューブリン複合体 (γ-tubulin complex) が局在する。それ以外にも、SPBと中心体を構成する因子には、互いに機能的な相同性をもつタンパク質が多く含まれる。 細胞分裂時には、 SPBは、 スピンドル微小管(高等生物の紡錘体:「酵母の紡錘体微小管」の項を参照)の形成起点として両極に存在し、微小管のマイナス端を繋ぎ止めて、精確な染色体の分配に貢献している。細胞分裂により母娘細胞に分配されたSPBは、1細胞周期に1回だけの複製を行って、細胞内での数を保っている。 酵母では、細胞分裂期にも核膜が消失しないため、SPBは、核膜に埋め込まれて、核内にスピンドル微小管を形成する(「酵母の核分裂」の項を参照)。 出芽酵母のSPBは、 複製されたばかりの新しいSPBが一過的に核膜の外に観察されるが、すぐに核膜に埋め込まれ、 その後は細胞周期を通して核膜に埋め込まれて存在する。分裂酵母のSPBは、分裂期にのみ核膜に埋め込まれて核内にスピンドル微小管を形成する。間期には、核膜に添うように細胞質側に存在し、細胞質微小管のMTOCのひとつとして働いている。 減数分裂では、染色体組換えの時期に、核内の微小管がSPBによってひとつに束ねられる。分裂酵母では、SPBが先頭になって核を引っ張りまわすように動き、相同染色体の効率的な組換えを可能にしている(「酵母の微小管」の項を参照)。 SPBと中心体は、MTOCとしての働きのほかにも、情報伝達分子の足場としての役割をもつ。分裂期に重要な複数のキナーゼが、 SPB・中心体に局在することが知られている。分裂酵母では隔壁形成を制御する分子群(SIN: septation initiation network)、出芽酵母では分裂期脱出を制御する分子群 (MEN: mitosis exit network)なども、SPBに局在する。複製されたSPBが等価ではない(成熟度の違いによってOldとNewの区別が生じる)ことを利用した、非対称や極性の制御に関わるしくみの存在が示唆される。細胞分裂に非対称性を与えることも、SPB・中心体の重要な役割であると考えられる。 参考文献 |

| 紡錘体形成 |
|---|
| 【Spindle assembly】 |
| 五島 剛太 |
| 名古屋大学大学院理学研究科生命理学専攻 |
| お問合せ |
紡錘体(スピンドル:spindle)は細胞分裂の際に形成される双極性(bipolar)の構造体であり、細胞分裂装置(mitotic apparatus)と呼ぶこともある。染色体を娘細胞へと分配し、細胞質分裂を制御するために必須の役割を果たす。主要構成因子はチューブリン(tubulin)の重合体である微小管(microtubule)、および、さまざまな微小管結合蛋白質(microtubule-associated protein: MAP)である。動物体細胞においては2つの中心体(centrosome)が主な微小管形成中心(microtubule organising centre: MTOC)となり、中心体から伸びた微小管が凝縮した染色体を捕捉する。一方、植物細胞や減数分裂期の動物細胞の一部には中心体が存在せず、別のメカニズムにより紡錘体微小管が生み出されている。これまでに、染色体自身が微小管の形成を誘導することや(Ran GTPアーゼやAurora Bキナーゼが関わる)、既存の微小管に依存して新たに微小管が生み出される微小管増幅経路(オーグミン複合体が関与する)の存在が明らかにされた。また、動物体細胞においてもこうした経路が中心体経路と同時に働いていることも明らかになった。参考文献 |
| 細胞分裂軸 |
|---|
| 【Cell division axis】 |
| 豊島 文子 |
| 京都大学ウイルス研究所 細胞生物学研究部門 構造形成学研究分野 |
| お問合せ |
| 細胞が分裂する方向のことであり、細胞分裂面と垂直に交わる軸方向を指す。細胞分裂軸は分裂期紡錘体の両極を結ぶ線と平行であるため、紡錘体軸の方向によって分裂軸が決定される。紡錘体軸は多くの場合、分裂中期に決定されるが、幾つかの組織・細胞では分裂後期に軸方向が変化するとの報告もある。
紡錘体軸の方向は「細胞の形」「細胞極性」「細胞―細胞間接着」「細胞ー細胞外基質接着」などの細胞内外の要因によって決定される。細胞は、周囲の環境や細胞の分化過程によって、これらの要因を使い分けている。そのため、紡錘体軸を決める分子機構は細胞や組織の種類によって異なる。進化的に保存されている紡錘体軸制御因子としてGα-LGN-NuMA複合体が広く知られている。しかし、全ての場合においてこの複合体が主要因子として機能するわけではないので、注意が必要である。 参考文献生化学 84, 81-91, 2012 |





