- ホーム
- >
- 細胞生物学用語集トップ
- >
- 細胞生物学用語集【あ行】
細胞生物学用語集【あ行】
【あ】
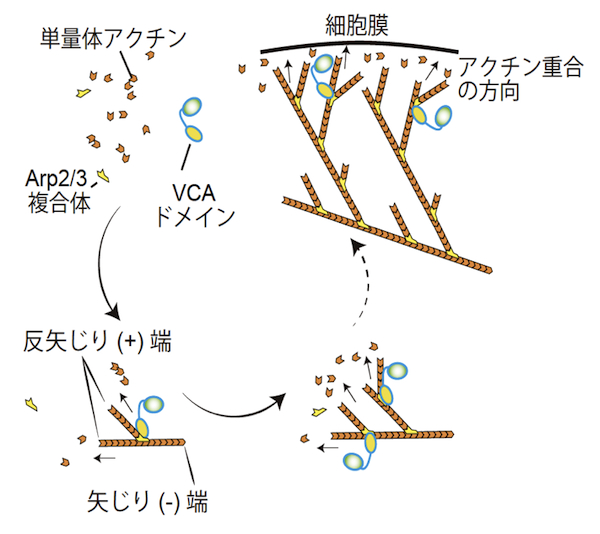
| Arp2/3複合体とアクチン重合 |
|---|
| 【Arp2/3 complex and actin polymerization】 |
| 千住 洋介・末次 志郎 |
| 東京大学 分子細胞生物学研究所 細胞形態研究分野 |
Arp2/3複合体は、二つのアクチン関連タンパク質 (actin-related protein) Arp2とArp3を含むヘテロ七量体のタンパク質複合体である。サブユニットのArp2とArp3は、ATP結合ポケットを持ち、単量体アクチン (G-actin) に似た構造を持つ。Arp2/3複合体は細胞に見られる枝分かれしたアクチン繊維の枝(branch)の基部に存在し、試験管内でも枝分かれしたアクチン繊維を形成することができる。この枝分かれ形成機構については、当初二つの説、end branchingおよびside branching、があったが、現在ではおおむね次のようなside branchingモデルに収束している。アクチンのみで行われるアクチン重合においては、単量体アクチンの3量体形成が重合核となる。したがって、Arp2/3複合体はアクチン重合に必要な重合核形成の段階で、単量体アクチンが形成する3量体のうち、2分子部分を代替すると考えられている。不活性化状態では、Arp2とArp3の配向はアクチン重合に適した方向ではないが、Arp2/3複合体とその活性化タンパク質との結合は、Arp2とArp3の配向をアクチン重合に適した方向に変える。代表的なArp2/3複合体の活性化タンパク質は、WASPファミリータンパク質であり、WASPファミリータンパク質のVCAドメインがArp2/3の活性化を担う。VCAドメインは、Arp2とArp3の配向を調節するだけでなく、3量体形成に必要な残り一つの単量体アクチンにも結合し、結合した単量体アクチンをArp2/3複合体に供給することでアクチン重合を活性化する。活性化したArp2/3複合体は、既に形成されているアクチン繊維 (F-actin) の側面に結合しつつ、アクチン重合を誘導し、アクチン繊維の枝分かれ構造を形成する【画像1】。このような枝分かれしたアクチン繊維の形成は、ラメリポディア形成やクラスリン被覆小胞のエンドサイトーシス等において重要であると考えられている。参考文献 |
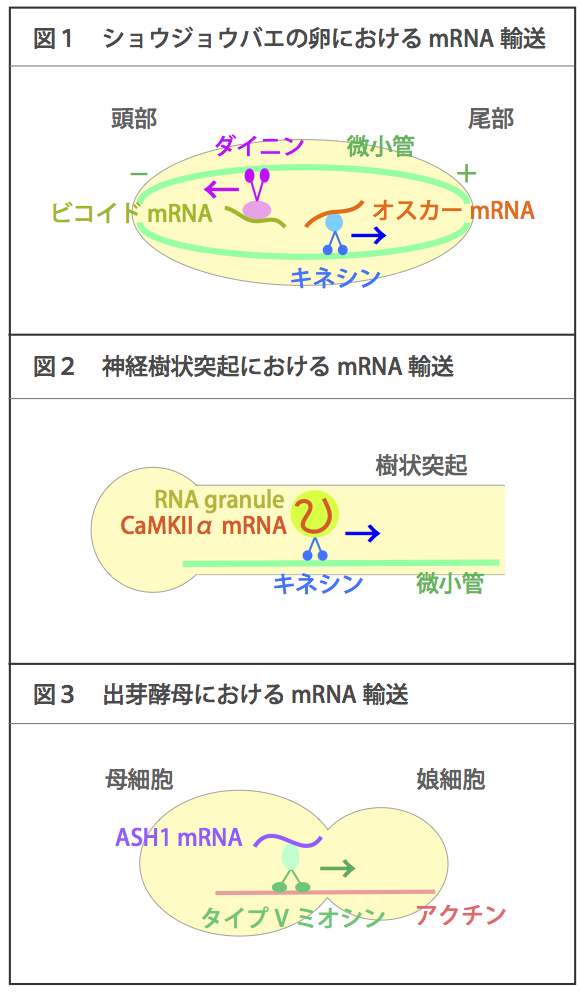
| RNAの輸送 |
|---|
| 【RNA transport】 |
| 椎名 伸之 |
| 基礎生物学研究所・神経細胞生物学研究室 |
| お問合せ |
| 微小管やアクチン細胞骨格をレールとして、特定の種類のmRNAが輸送される。微小管をレールとした場合にはキネシンやダイニンをモータータンパク質として用い、アクチンをレールとした場合にはタイプVミオシンをモータータンパク質として用いることが知られている。mRNA輸送によって、細胞内の局所でタンパク質を合成して機能させることができる。 例えばショウジョウバエの卵では、微小管のマイナス端は頭部側を、プラス端は尾部側を向いて形成されている。ビコイドmRNAはダイニンによってマイナス端すなわち頭部側へ輸送され、オスカーmRNAはキネシンによってプラス端すなわち尾部側へ輸送される(図1)。頭部、尾部にそれぞれ係留されたmRNAから局所的にタンパク質が合成される結果、タンパク質局在に濃度勾配ができる。その濃度勾配は、頭部と尾部の体軸形成に必須である。 神経細胞の樹状突起では、カルモジュリンキナーゼIIα(CaMKIIα) mRNAなどが微小管をレールとしてキネシンによって輸送される。この際mRNAはRNA granuleと呼ばれる巨大複合体に取り込まれて輸送されることが知られている(図2)。mRNAはシナプス刺激入力に応じて局所的に翻訳され、局所シナプスの選択的な長期増強、ひいては記憶や学習に関与する。 また、出芽酵母では、ASH1 mRNAがアクチンをレールとしてタイプVミオシンによって母細胞から娘細胞へ輸送される(図3)。その結果、ASH1 mRNAおよび翻訳産物は娘細胞にのみ局在し、母細胞と娘細胞の運命(接合型)は異なったものになる。 参考文献 |
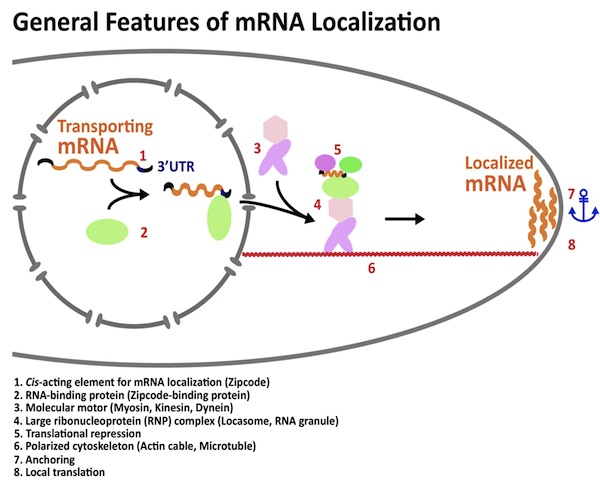
| RNA局在 |
|---|
| 【RNA localization 】 |
| 入江 賢児 |
| 筑波大学大学院人間総合科学研究科(基礎医学系)分子細胞生物学グループ |
| お問合せ |
| 「RNAの局在化」とは、核内で転写されたRNAが細胞内の特定の場所に存在する現象です。RNAの局在化により、細胞内では情報の偏りが生じ、細胞の極性が生まれます。生物の遺伝情報は、遺伝子の本体であるDNAからRNAに写し取られて発現します。そのため、RNAの局在化は、遺伝子発現を空間的に制御するための重要な現象です。図の説明 (1) 局在化するmRNAはジップコードと呼ばれる局在化シグナルをもっている。ジップコードはmRNAの3’非翻訳領域に見られることが多い。
(2) ジップコードにRNA結合タンパク質が結合する。 (3) ミオシンやキネシンなどの分子モーターがRNA局在に使われる。 (4) mRNAとRNA結合タンパク質は分子モーターとともに、巨大なRNA-タンパク質複合体を形成する。 (5) 輸送されるmRNAの翻訳は抑制されることが多いので、巨大なRNA-タンパク質複合体には翻訳の抑制因子が含まれる。 (6) RNA-タンパク質複合体はアクチンケーブルや微小管などの極性化した(方向性をもった)細胞骨格上を輸送される。 (7)と(8) 輸送されるmRNAは局在化部位に係留され、そこで局所的に翻訳される。 参考文献1;Moving messages: the intracellular localization of mRNAs. St Johnston D. Nat Rev Mol Cell Biol. 2005 May 6(5):363-75. |
| RNA制御 |
|---|
| 【RNA regulation】 |
| 入江 賢児 |
| 筑波大学大学院人間総合科学研究科(基礎医学系)分子細胞生物学グループ |
| お問合せ |
遺伝情報はDNAからRNAを経てタンパク質として発現される。 生物の持つ複雑で巧妙な形態・機能の獲得には、RNA段階での 遺伝子発現制御プログラムが重要な役割を果たす。すなわち、 個体発生の過程において、様々な「1.非対称性制御プログラム」 により、単一の受精卵から非対称な細胞群が生成され、「2.多様 性獲得プログラム」(選択的スプライシング)により、分化過程で 形成される細胞が担う多様な機能の獲得に必要な遺伝子産物 自体の多様性が獲得される。さらに、「3.品質保証プログラム」 による厳密な監視により、RNAレベルでの制御の正確性が保証される。
参考文献1;蛋白質核酸酵素 2009年12月増刊号 mRNAプログラム。 |
| アクチン重合 |
|---|
| 【actin polymerization】 |
| 渡邊 直樹 |
| 東北大学 大学院生命科学研究科 |
| お問合せ |
アクチンは筋肉の線維性タンパク質であるが、筋細胞以外のほとんどの真核細胞にも存在し、表層の細胞骨格ネットワークの主成分を成している。アクチンは単量体、もしくは、それが数珠状につながった線維のいずれかの状態で存在する。細胞が運動するときや外来の刺激に応答し変形する際、単量体アクチンが線維に変換するアクチン重合が盛んにおきる。アクチン重合は、線維を形成する重合核形成(nucleation)と線維端に単量体が次々と付加する伸長(elongation)の2つのステップに分けることができる。重合核形成のステップでは、単量体アクチンが2量体や3量体を形成しては解離することを繰り返すため、なかなか線維が形成されない。この重合核形成を促進する分子群として、Arp2/3複合体、フォルミンファミリータンパク質、SpireなどWH2ドメインをもつものが同定されてきた。これらの重合核形成促進因子は、Rhoファミリー低分子量Gタンパク質を中心とした細胞内情報伝達機構によって活性化される。アクチン伸長は、フォルミンファミリーやVASPによって維持、もしくは加速され、逆にキャッピングプロテイン、ゲルソリンなどによって阻害される。これらの機構により細胞内のアクチン重合のタイミングは調整され、細胞表層の構造をリモデリングするとともに、細胞先端の仮足では重合端を外側に向けながら細胞膜を押す力を発生する。参考文献 |
| アクチン繊維によるメカノセンシング |
|---|
| 【Mechanosensing by actin filaments】 |
| 上田 太郎 |
| 産業技術総合研究所 バイオメディカル研究部門 |
| お問合せ |
| アクチン繊維は弾性をもった二重らせんであり、引っ張ればわずかに長くなる[1,2,3]。Shimozawaら[4]は、蛍光標識したアクチンの蛍光強度がin vitroの張力負荷によって変化することを見出しているし、Matsushitaら[5]は、分子動力学シミュレーションにより、張力負荷がアクチン繊維の長さとらせんピッチ両方を伸ばすことを報告している。したがって張力負荷されたアクチン繊維の長さとらせんピッチがばねのように伸びることは間違いない。しかしこのことが直ちにアクチン繊維がメカノセンサーであることを意味するわけではない。アクチン繊維がメカノセンサーとして機能するためには、張力負荷によりアクチン繊維としての機能が変化し、何らかのシグナル伝達の起点になければならないからである。しかしアクチン繊維は弾性係数が大きく(バネとして固い)、生理的な張力負荷で伸びる割合が小さい(0.3%程度)ため、これがシグナル伝達の起点になるという考えは一般的ではなかった。
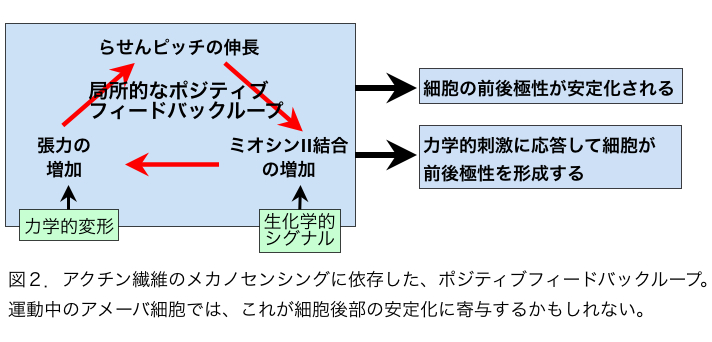
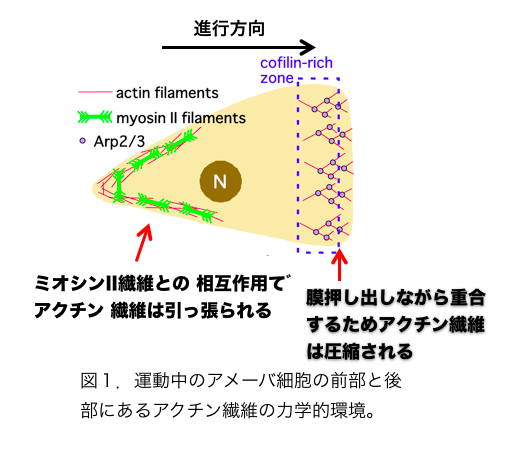
こうした状況にあって、2011年にアクチン繊維はメカノセンサーであることを示す論文が2報、わが国の研究者から発表された。論文発表は同じ2011年でも学会発表が先行していたという意味で第一にあげるべきは、名古屋大の早川、辰巳、曽我部らによる、コフィリンとアクチン繊維の相互作用に関する論文である[6]。彼らはin vitroの実験で、コフィリンは張力のかかったアクチン繊維と結合しにくく、また切断しにくいことを発見し、さらにセミインタクトおよびインタクトなhuman umbilical vein endothelial cells (HUVECs)において、コフィリンが張力のかかっていないストレスファイバーと選択的に結合することも示した。張力のかかっていないストレスファイバーが崩壊しやすいことは以前から知られており、アクチン繊維のメカノセンシングに依存したコフィリン活性がこの現象に関与すると示唆されたわけである。もう一つの例は張力に依存したアクチン繊維とS1(ミオシンIIのモータードメイン)の結合である[7]。これはもっぱら生きた細胞性粘菌細胞を用いた観察で、GFP−S1が張力がかかっている細胞内アクチン繊維と優先的に結合することを報告ている。この現象は、S1にアクチンとの親和性を高める変異が必要であること、またミオシンIIとアクチン繊維の相互作用を調節することが知られている既知のタンパク質に依存しないことなどから、張力のかかったアクチン繊維に特徴的な構造をS1が直接認識して優先的に結合したと解釈された。興味深いことに、ミオシンIIのS1は張力負荷アクチン繊維と優先的に結合したが、ミオシンIのモータードメインはそうした選択性を示さなかった。この現象は、ミオシンIIは本来、細胞後部でフィラメントを形成してアクチン繊維と相互作用し、張力を発生させるのに対し、ミオシンIは細胞前部に局在して張力発生には関与しないことと関連があるのかもしれない。現在のところ、アクチン繊維のメカノセンシングが関与することが示唆されているのは、コフィリンおよびミオシンIIとアクチン繊維の相互作用だけであるが、アクチン繊維の構造が張力負荷で変化するなら、他のアクチン結合タンパク質との相互作用もアクチン繊維のメカノセンシングにより様々な調節を受けていると推測される。 アクチン系細胞骨格と相互作用するメカノセンサーはすでにいくつか知られている[8,9,10,11,12]。一方、アクチン繊維は非常に多数の因子と相互作用する多機能構造であり、かつ細胞表層全体に張り巡らされたネットワークであるため、アクチン繊維自体がメカノセンサーであることには、細胞全体の活性をコーディネートしうるという意味で大きな生理的意義をもちうる。こうした観点から議論しておきたいことは、アクチン繊維の構造多型と機能分化の関連である。そもそもアクチン繊維が単体で構造的多型性をもつという概念はE.H.Egelmanらの電子顕微鏡観察によりかなり以前から提唱されていた[13,14,15] (ただし最近のFujiiらの電子顕微鏡観察[16]はアクチン繊維の構造多型性に否定的であり、決着はついていない)が、その生理的意義は不明確であった。しかしコフィリンを初めとする種々のアクチン結合タンパク質の結合が、アクチンサブユニットの構造を変化させることが明らかとなり[17]、これがアクチン結合タンパク質とアクチン繊維の結合が協同的であることの構造的基盤であると提唱され[18]、アクチン繊維の構造多型の生理的意義がおぼろげながら見えてきた。つまり、たとえばコフィリンがアクチン繊維のあるサブユニットに結合すると、そのサブユニットの構造が「コフィリン結合型」に変化し、さらにこの構造変化が隣接するサブユニットの構造もコフィリン結合型に変化させる(協同的構造変化)と、その隣接サブユニットとコフィリンの親和性が増加し、結果的にコフィリンのクラスターが形成される(協同的結合)という考え方である。一方、ミオシンIIがアクチン繊維に結合すると、張力とは無関係にアクチン繊維のらせんピッチがわずかに延びることが報告されており[19]、またHMM(ミオシンIIの可溶性断片)は、微量のATP存在下でアクチン繊維と協同的に結合することも報告されている[20]ので、ミオシンIIモーターとアクチン繊維の協同的相互作用にもコフィリンの場合と同様、アクチン繊維の協同的構造変化が関与する可能性がある。ただしらせんピッチの伸び縮みの方向が逆なので、コフィリン結合によるアクチン繊維の協同的構造変化と、ミオシンII結合によるアクチン繊維の協同的構造変化はまったく異なったものであろう。この仮想的メカニズムには、細胞内のアクチン繊維の機能分化を安定化させる作用が期待でき、大変魅力的である(図1,2)。 参考文献1. Kojima H, Ishijima A, Yanagida T (1994) Proc Natl Acad Sci U S A 91: 12962-12966.
2. Huxley HE, Stewart A, Sosa H, Irving T (1994) Biophys J 67: 2411-2421 3. Wakabayashi K, Sugimoto Y, Tanaka H, Ueno Y, Takezawa Y, et al. (1994) Biophys J 67: 2422-2435. 4. Shimozawa T, Ishiwata S (2009) Biophys J 96: 1036-1044. 5. Matsushita S, Inoue Y, Hojo M, Sokabe M, Adachi T (2011) J Biomech 44: 1776-1781. 6. Hayakawa K, Tatsumi H, Sokabe M (2011) J Cell Biol 195: 721-727. 7. Uyeda TQ, Iwadate Y, Umeki N, Nagasaki A, Yumura S PLoS One 6: e26200. 8. Yonemura S, Wada Y, Watanabe T, Nagafuchi A, Shibata M (2010) Nat Cell Biol 12: 533-542. 9. Sawada Y, Tamada M, Dubin-Thaler BJ, Cherniavskaya O, Sakai R, et al. (2006) Cell 127: 1015-1026. 10. Kee YS, Robinson DN (2008) Curr Biol 18: R860-862. 11. Pramanik MK, Iijima M, Iwadate Y, Yumura S (2009) Genes Cells 14: 821-834. 12. del Rio A, Perez-Jimenez R, Liu R, Roca-Cusachs P, Fernandez JM, et al. (2009) Science 323: 638-641. 13. Egelman EH, Francis N, DeRosier DJ (1982) Nature 298: 131-135. 14. Galkin VE, Orlova A, Schroder GF, Egelman EH (2010) Nat Struct Mol Biol 17: 1318-1323. 15. Orlova A, Prochniewicz E, Egelman EH (1995) J Mol Biol 245: 598-607. 16. Fujii T, Iwane AH, Yanagida T, Namba K (2010) Nature 467: 724-728. 17. Galkin VE, Orlova A, Lukoyanova N, Wriggers W, Egelman EH (2001) J Cell Biol 153: 75-86. 18. Ressad F, Didry D, Xia GX, Hong Y, Chua NH, et al. (1998) J Biol Chem 273: 20894-20902. 19. Tsaturyan AK, Koubassova N, Ferenczi MA, Narayanan T, Roessle M, et al. (2005) Biophys J 88: 1902-1910. 20. Tokuraku K, Kurogi R, Toya R, Uyeda TQP (2008) J Mol Biol 386: 149-162. |
【い】
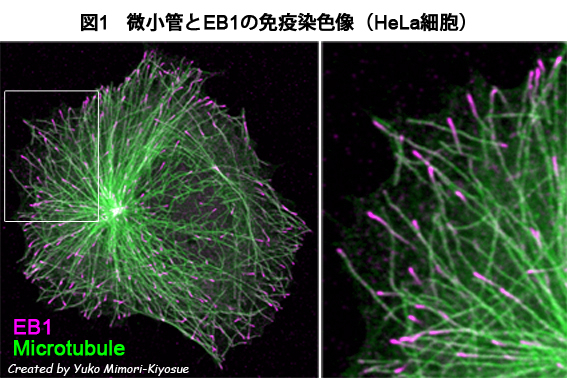
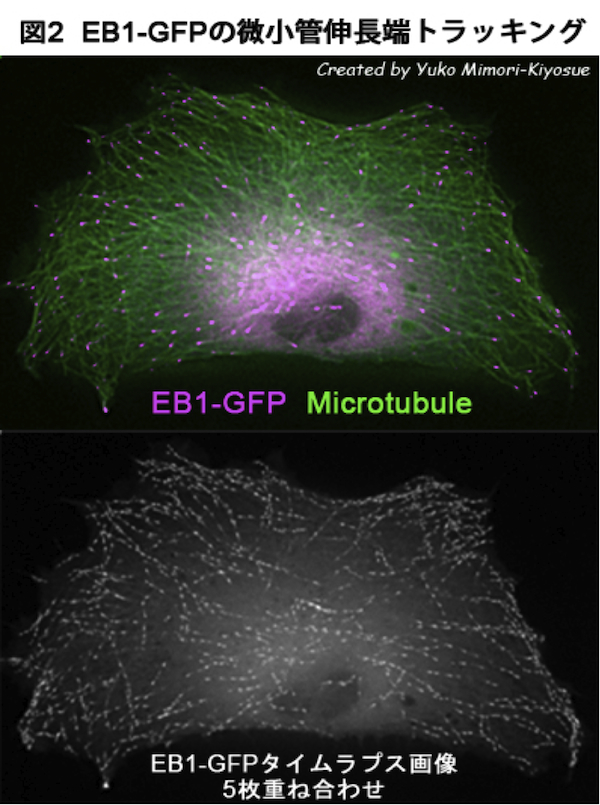
| EB1ファミリー・タンパク質 |
|---|
| 【EB1 family proteins】 |
| 清末 優子 |
| 理化学研究所 神戸研究所 発生・再生科学総合研究センター (CDB) |
| お問合せ |
| EGFRトランスアクチベーション |
|---|
| 【EGFR transactivation 】 |
| 目加田 英輔 |
| 大阪大学微生物病研究所細胞機能分野 |
リゾフォスファチジン酸やエンドセリン、アンジオテンシンIIなどによるG蛋白共役型受容体(GPCR)の活性化に伴って、EGFRが活性化される現象をEGFRトランスアクチベーションと呼ぶ。この過程でHB-EGFのシェディングが中心的な役割を果たしていることが近年明らかになってきている。GPCRリガンドによるGPCRの活性化がRas-ERK経路を活性化し、このことがHB-EGFのシェディングを誘導し、分泌されたHB-EGFがEGFRを活性化し、さらにその下流でRas-ERK経路が活性化する。このことがさらなるHB-EGFのシェディングのみならずHB-EGF自身の発現をも誘導し、ここに細胞増殖あるいは移動促進などシグナルのポジティブ・フィードバックループが形成される。このようなEGFRトランスアクチベーション機構とこれによるポジティブループの形成が、心肥大や動脈硬化、高血圧、そして発癌など様々な病理にも深く関わっていることが近年わかってきている。参考文献蛋白質核酸酵素 54, 1722-1727, 2009 |

| EGFファミリー |
|---|
| 【EGF family 】 |
| 目加田 英輔 |
| 大阪大学微生物病研究所細胞機能分野 |
EGFファミリーの増殖因子はEGFRに結合する。一方、EGFRにもErbB2, ErbB3, ErbB4のファミリー分子が存在する。Neuregulin (NRG)はEGFRには結合せずErbB3, ErbB4に結合する。各ErbBはお互いにホモあるいはヘテロダイマーを形成して機能する。故に、例えばHB-EGFの場合、HB-EGFはEGFRとErbB4に結合するのでEGFRとErbB4を含む全てのホモあるいはヘテロダイマーを活性化しうる。このようなリガンド-受容体の対応関係が、受容体下流のシグナルの多様性を現すひとつの理由と考えられる。狭義にEGFファミリーという場合はEGFRに結合する増殖因子を指すことが多いが、広義にはNRG等ErbBファミリーに結合する増殖因子も含める。EGFファミリーの特徴として、これらの分子が膜型前駆体蛋白質として合成された後にエクトドメイン・シェディングと呼ばれる酵素的切断を受けて分泌されることがあげられる。参考文献Cytokine Growth Factor Rev. 11: 335-344. 2000 |

| ERMファミリー |
|---|
| 【ERM family】 |
| 田村 淳・月田 早智子 |
| 大阪大学大学院生命機能研究科 |
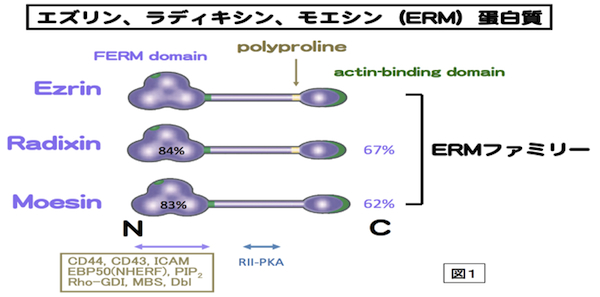
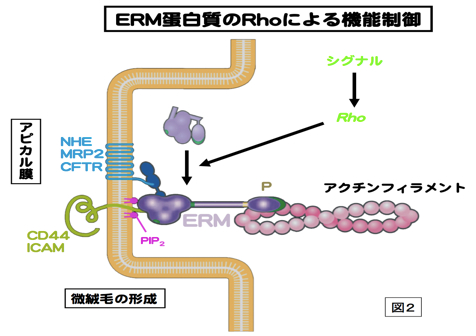
細胞骨格の1つであるアクチンフィラメントと細胞膜との動的な相互作用をつかさどる蛋白質ファミリー。微絨毛や神経成長錐、細胞分裂時の収縮環などアクチンフィラメントが細胞膜に結合する部分に局在する。アミノ酸レベルで75%程度の相同性を持つ3つの蛋白質Ezrin,Moesin,RadixinによりERMファミリーが形成され、各々の構成蛋白質はERM蛋白質とよばれる。いずれもN末側からFERMドメイン、α-ヘリックス領域、アクチン結合モチーフの順に並ぶドメインを持つ、分子量75kDaほどの分子である(図1)。エズリンとラディキシンは、モエシンにはないポリプロリンモチーフを余分に持つ。ERM蛋白質は、細胞膜-アクチンフィラメントを架橋する棒状の活性型と球状の不活性型の2つのコンフォメーションをとる。ERM蛋白質はRhoの下流で、PIP2により活性化されて、細胞膜とアクチンフィラメントに、各々、N末のFERMドメインとC末で結合すると考えられる。活性化型ERM蛋白質はC末のトレオニン残基のリン酸化により、安定化される。細胞膜との結合では、CD44などの細胞膜蛋白質に直接結合する場合と、トランスポーターなどの細胞膜蛋白質にPDZドメインをもつEBP-50を介して結合する場合がある(図2)。各々のERM蛋白質は、培養細胞では共発現することが多く、同じ局在を示すことも多いが、生体組織レベルでは、組織の種類によって異なる発現と局在を示す。例えば、肝細胞ではRadixinの発現が多く、腸管上皮細胞ではEzrinが、血管内皮細胞ではMoesinが多く発現している。Radixin、Ezrinのノックアウトマウスでは各々肝臓および胃の上皮細胞において、MRP2やHK-ATPaseのアピカル細胞膜への局在不全のためにDubin-Johnson型の高ビリルビン血症や胃酸分泌障害になる。上皮細胞アピカル表層構築に重要な蛋白質として、注目されている。参考文献 Tsukita, S., Yonemura, S. (1999)
Kikuchi, S., Hata, M., Fukumoto, K., Yamane, Y., Matsui, T., Tamura, A., Yonemura, S., Yamagishi, H., Keppler, D., Tsukita, S., Tsukita, S. (2002) Fehon, RG., McClatchey, AI., Bretscher, A. (2010) Arpin, M., Chirivino, D., Naba, A., Zwaenepoel, I. (2011) Neisch, AL., Fehon, RG. (2011) |
| 1分子イメージング法 |
|---|
| 【Single-molecule imaging technique】 |
| 原田 慶恵 |
| 京都大学 物質-細胞統合システム拠点(iCeMS) |
| お問合せ |
| 生体分子に様々な標識を付けて、それを手がかりに光学顕微鏡で個々の分子の局在や運動等を観察する手法を1分子イメージングあるいは1分子観察という。標識は大きく分けて2種類ある。大きな標識と、蛍光を発する標識である(蛍光1分子イメージング法の項を参照)。大きな標識を使う場合、1分子イメージングに特別な装置は不要で、光学顕微鏡で容易に観察できるという利点がある一方で、生体分子1個の観察ができているか、機能が損なわれていないかについての注意が必要である。大きな標識として直径1μm程度のマイクロビーズがよく使われる。マイクロビーズは光ピンセット(1分子操作法の項参照)で操作することや位置を精密に計測する実験(1分子操作法の項参照)にも使うことができ、1分子イメージングと1分子操作、1分子計測を組み合わせた実験を行うことができる。 |
| 1分子計測法 |
|---|
| 【Single-molecule measurement technique】 |
| 原田 慶恵 |
| 京都大学 物質-細胞統合システム拠点(iCeMS) |
| お問合せ |
| 生体分子に結合させた標識の位置や動きを高精度で1分子計測することで、個々の分子がどのようなメカニズムで機能しているのかを明らかにすることができる。
マイクロビーズなどの大きな標識は可視光で照明し、対物レンズでその像(影)を大きく(1000から10万倍)拡大し4分割のフォトダイオードに投影することで、その動きを1nm以下の精度で検出することができる。ただし、微小な動きを検出する場合は、標識のブラウン運動を防ぐために光ピンセット等で捕捉する必要がある。また、光ピンセットで捕捉することによって、そのバネ定数と検出した変位から、ビーズにかかっている力を見積もることができる。モータータンパク質であるキネシン分子を固定した直径1μmのビーズを光ピンセット(バネ定数~0.1pN/nm)で捕捉し微小管と相互作用させることで、キネシン分子が微小管に沿って8nmずつステップ状に動くことと最大7pNの力を出すことが明らかになった。また、RNAポリメラーゼがDNAの情報を1塩基ずつ読み進んでいく0.34nmステップの動きも同様の方法で検出された。 蛍光色素分子や量子ドットなどを1分子イメージング蛍光顕微鏡で観察した場合、直径が数百nmのぼやけた像として観察されるため、そのままでは、正確な位置はわからない。しかし、この像の光強度のプロファイルを解析することによってその中心点、すなわち蛍光物質の存在している位置をnm精度で決定することができる。この方法を使って二量体のモータータンパク質分子であるミオシンVが2つのモータードメインを交互に動かすことによってアクチンフィラメントに沿って運動することが明らかになった。 最近は量子ドットを使って、細胞内で1分子計測が行われ始めている。 |
| 1分子操作法 |
|---|
| 【Single-molecule manipulation technique】 |
| 原田 慶恵 |
| 京都大学 物質-細胞統合システム拠点(iCeMS) |
| お問合せ |
| 生体分子の機能を調べるためには個々の分子を操作する手法が有効である。1分子を捕捉、操作する方法としてレーザー光を対物レンズで集光させ、直径1~数μm程度の微小粒子を光でトラップする「光ピンセット」、プラスチックの中に酸化鉄が封入された磁気ビーズや磁性粒子を磁石を使って操作する「磁気ピンセット」、原子間力顕微鏡(AFM)の力測定モードなどが使われる。光ピンセットや磁気ピンセットの最大捕捉力は100pN程度、AFMは固いカンチレバーを使うことで、数百pN以上の大きな力で捕捉することができる。 2つの光ピンセットを使って、2個のビーズを捕捉し、その間に1本のDNA分子を伸展させ、RNAポリメラーゼ分子の結合を観察する実験や、DNAの片端をガラス基板上固定し、もう一方の端には磁気ビーズに結合させ、磁気ピンセットでDNAをねじった後、磁気ビーズの高さ変化を計測することで、トポイソメラーゼがATPを加水分解し、ねじれを解消する反応を検出した実験など、光ピンセットや磁気ピンセットはDNAを使った実験に使われることが多い。また、AFMを使い、カンチレバーの先端にタンパク質分子を結合させ、それを引っ張ってタンパク質分子のドメイン構造が壊れていく様子などが観察されている |
【え】
| FCMR/FAIM3/TOSO |
|---|
| 【FCMR/FAIM3/TOSO 】 |
| 大野 博司 |
| 理化学研究所・横浜研究所 免疫アレルギー科学総合研究センター(RCAI) |
| お問合せ |
FCMR/FAIM3/TOSOはFas仲介型のアポトーシスを阻害する因子として発見された分子である。その後、他のFc受容体とのアミノ酸配列の相同性から、IgMと結合する受容体として同定された(1)。PreB細胞以降のB細胞の成熟過程で発現していること(参考文献1, 図1)や、慢性リンパ球性白血病での発現量の増加(参考文献2)その発現がB細胞のシグナリングによって調節されていること(参考文献3)などから、B細胞の活性化や、シグナリングに影響を与えていることが予測される。参考文献(1)Shima, et al. Int. Immunol. (2010)
(2)Proto-Siqueira, et al. Blood (2008) (3) Pallasch, et al. Blood (2008) 抗体 |

| M-Sec/TNFaip2/B94/EXOC3L3 |
|---|
| 【M-Sec/TNFaip2/B94/EXOC3L3】 |
| 大野 博司 |
| 理化学研究所・横浜研究所 免疫アレルギー科学総合研究センター(RCAI) |
| お問合せ |
正常ヒト臍帯静脈内皮細胞(HUVEC)をTNFα処理することによって発現上昇する遺伝子として始めに単離された、Sec6ファミリーに属する細胞質蛋白質。発生段階における造血系組織、精子の先体に高く発現する。低分子GTP結合蛋白質RalAとエキソシスト複合体と協調して、膜ナノチューブ形成に関わる。参考文献Nat. Cell. Biol. 11 (12), 1427-32, J. Biol. Chem. 269(5), 3633-40, |
| M細胞、GP2 |
|---|
| 【Microfold cell (M cell)、Glycoprotein2 (GP2) 】 |
| 大野 博司 |
| 理化学研究所・横浜研究所 免疫アレルギー科学総合研究センター(RCAI) |
| お問合せ |
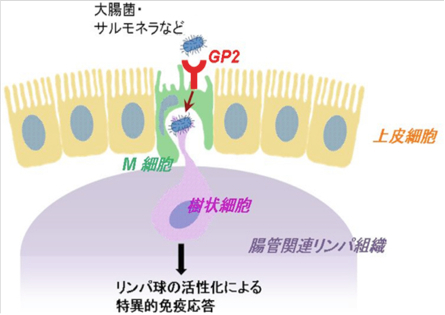
| Microfold cell (M cell)はパイエル板を始めとする粘膜関連リンパ組織を覆う特殊な上皮層に点在する抗原取り込み細胞である。M細胞は周囲の腸管吸収上皮細胞とは形状が異なり、微絨毛が短く疎で、基底膜側は免疫細胞を抱え込むための陥凹した空間を持つ(図1)。このような特徴により粘膜組織に侵入した外来抗原を免疫細胞へと効率よく受け渡すことができる。
Glycoprotein2(以下GP2)はGPI-アンカー型タンパク質の一つで、膵臓腺房細胞に限局して発現する機能未知の分子として同定された(文献1)。その後の研究からマウス、ヒトのM細胞の頂端面細胞膜に発現することが見出され、1型線毛の先端に表出するFimHタンパク質を認識することが明らかとなった(文献2)。M細胞においてGP2は1型線毛を持つ細菌を選択して認識し、細胞内へと取り込むための受容体であり、粘膜免疫にける免疫監視において重要な役割を担う分子である(図1)。 参考文献文献1. Fukuoka S, et al. Proc Natl Acad Sci U S A. 1992 Feb 15 89(4):1189-93. 文献2. Hase K, et al. Nature. 2009 Nov 12 462(7270):226-30. 理化学研究所、免疫系構築研究チーム 抗体情報:販売元MBL Anti-GP2 / Glycoprotein2Anti-GP2 / Glycoprotein2 (Mouse) |
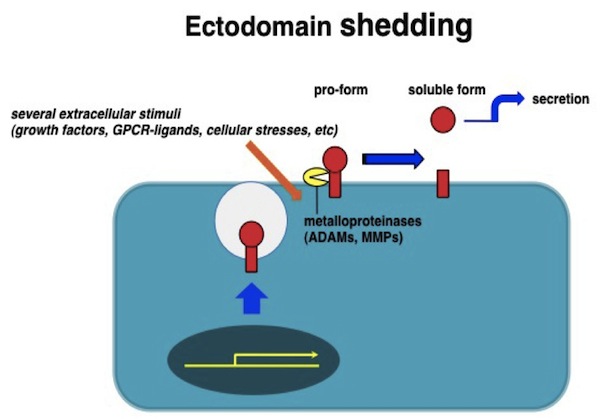
| エクトドメイン・シェディング |
|---|
| 【Ectodomain shedding 】 |
| 目加田 英輔 |
| 大阪大学微生物病研究所細胞機能分野 |
細胞増殖因子やサイトカイン、またこれらに対する受容体、あるいは細胞接着因子や細胞外マトリックスなど様々な細胞間情報伝達を担う分子群のある種のものは、細胞外からの刺激に伴って酵素的な切断を受け、細胞外に放出されることが古くから知られている。このプロセスはエクトドメイン・シェディングと呼ばれ、膜蛋白質を細胞膜にアンカーされた状態から細胞外に遊離させることで、分子の存在状態を劇的に変化させる。EGFファミリーの増殖因子をはじめとするある種の増殖因子やサイトカインは、はじめ膜結合型の前駆体蛋白として合成された後、シェディングによって細胞外に分泌される。エクトドメインシェディングは、特に膜結合型として合成される増殖因子やサイトカインにおいて情報の発信を制御する重要な機構のひとつである。種々のHB-EGF変異マウスの解析から、マウス生体内においてHB-EGFが正常にその機能を発揮するために、エクトドメインシェディングが非常に重要なステップであることが明らかになっている。 参考文献蛋白質核酸酵素 54, 1722-1727, 2009 |