- ホーム
- >
- 細胞生物学用語集トップ
- >
- 細胞生物学用語集【は行】
細胞生物学用語集【は行】
【は】
| BAR ドメイン |
|---|
| 【BAR domain】 |
| 千住 洋介・末次 志郎 |
| 東京大学 分子細胞生物学研究所 細胞形態研究分野 |
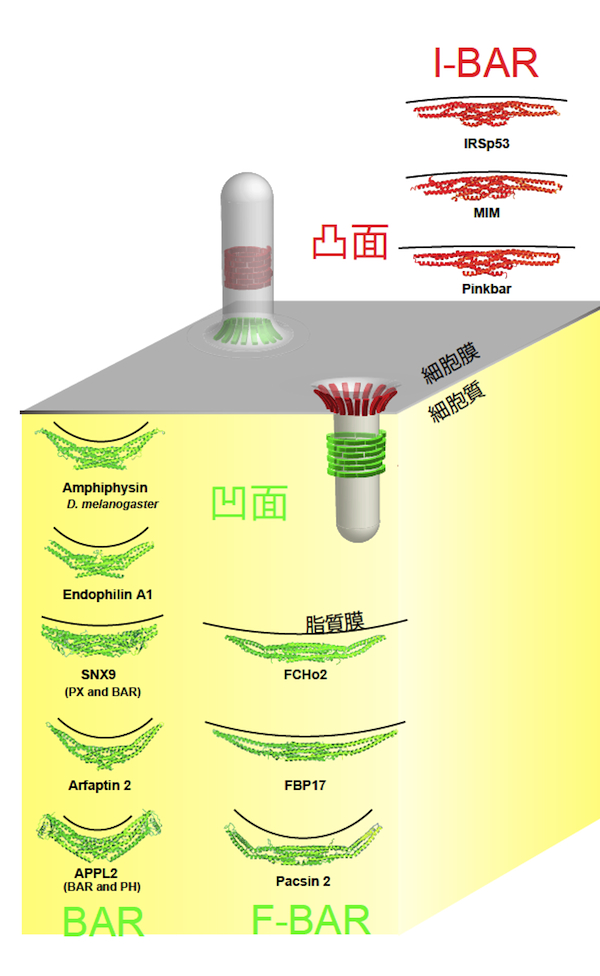
BAR (Bin/Amphiphysin/Rvs) ドメインスーパーファミリーは、BARドメインと総称される、BARドメイン、F-BARドメイン、I-BARドメインのサブファミリーに分類されるドメインを持つ。BARドメインおよびF-BARドメインは、三日月形の二量体を形成し、負に帯電している細胞膜と静電的な相互作用で結合する【画像1】。BARドメインとF-BARドメインは、三日月型二量体の立体構造における凹面が正に帯電している。この結合様式は、細胞膜の陥入構造に対応すると考えられ、実際に、多くのBARドメイン含有タンパク質 (Amphiphysin やEndophilinなど) や、F-BARドメイン含有タンパク質 (FBP17やFCHo1など) は、エンドサイトーシスなどの細胞膜の陥入構造の形成に関与する。一方、IRSp53等に含まれるI-BARドメインは、凸面が正に帯電している。よって、陥入構造とは逆の形態である、フィロポディアなどの細胞膜の突起構造の形成に関与する。また、BARドメイン、F-BARドメイン、I-BARドメインの中には、両親媒性ヘリックスなどの脂質膜に挿入されると考えられる部位を持つものがあり、この部位の挿入は、細胞膜の曲率形成を促進する。全体として、BARドメインスーパーファミリーは、細胞膜の曲率を認識、あるいは生成する機能を有すると考えられている。多くのBARドメイン含有タンパク質は、SH3ドメイン等の他のドメインを伴う。このため、BARドメイン含有タンパク質は、膜の曲率に準拠して、タンパク質を集積させる可能性がある。代表的なSH3ドメイン結合タンパク質は、Arp2/3複合体を活性化するWASPファミリータンパク質や、細胞膜を切断するダイナミン (dynamin) である。参考文献 |
| 培養基質の制御と細胞 |
|---|
| 【Regulation of culture substrate and cellular function】 |
| 原田 伊知郎 |
| 東京工業大学大学院生命理工学研究科 |
| 臓器・組織から取り出した細胞は培養液中で浮遊もしくはプラスチックシャーレの底に接着させて培養するのが一般的である。接着培養する細胞は、シャーレの底に物理吸着した細胞外マトリクス(以下ECM: Extracellular matrix) にインテグリン(Integrin)という膜タンパク質を介して接着している。したがって、細胞とECMとの接着の分子機構自体は生理的なものである。しかし、細胞外マトリクスが吸着しているプラスチックシャーレが生理条件下ではあり得ないほどの固いため、その環境は生体内とは著しく異なっていると考えられている。このような接着している細胞の足場(scaffold)の固さが細胞の機能に直接影響を及ぼしているのではないかという配慮から、近年様々な柔らかい培養基質が考案されている。 培養基質の固さの制御方法として細胞生物学的基礎研究によく用いられているのは電気泳動に用いるアクリルアミドゲルを培養基質とする方法である。アクリルアミドゲルは作成する際に添加する架橋剤の濃度を調整するだけで、容易に様々な固さのものを作成することができる。しかし、このゲルはタンパク質がほとんど吸着しないため、細胞外マトリクスもほとんど吸着しない。そのため、方法は様々であるがアクリルアミドゲルとECMとを架橋剤を介して結合させるのが一般的である。この培養基質を用いた研究によって、通常シャーレに培養した細胞に見られる極端な細胞骨格の発達や、アメーバのように広がった細胞の形はシャーレの固さが原因であったことが示されつつある。また、培養基質の固さを調整することで、細胞の分化能や増殖能などの機能も制御できることが示されている。 他にも様々な培養基質が考案されているものの、入手しやすいアクリルアミドゲルが多く用いられているもう一つの理由として、ゲルの透明度が上げられる。そのため細胞が観察しやすく広く普及している。特に、このゲル中に直径0.2〜1μmのマイクロビーズを包埋しておくと、ビーズの動きからゲルの歪みを計測することが出来るため、細胞が足場に力をかけている様子までも観察できる。そのため、細胞は移動に伴いどのような力を足場にかけているのか、等のような基礎研究にも本培養基質が用いられており、現在はTraction Force Microscopyとよばれ、新しい解析手法として普及している。 細胞が足場に加える力の制御・計測する他の方法として、ECMのマイクロパターニングやマイクロピラーを用いた研究も報告されている。カバーガラスにECMをプリントし、その形に細胞の形態を制御することで細胞骨格の発達を抑制し、柔らかい足場と同等の効果をねらった研究が報告されている。また、アクリルアミドゲルを用いたTraction Force Microscopyでは、接着点に直接かかっている力の定量化は逆問題であることから厳密な解析は難しい。そこで、3次元的なマイクロパターニング技術により、マイクロピラーをシリコンゴム(PDMS: polydimethylsiloxane)で作成して培養基板とすることで、一つ一つの接着点にかかる力のイメージングも試みられている。 参考文献(1) Pelham, R.J., and Y.L. Wang. 1997. Cell locomotion and focal adhesions are regulated by substrate flexibility. Proc Natl Acad Sci USA. 94:13661-13665.
(2) Munevar, S., Y. Wang , and M. Dembo. 2001. Traction force microscopy of migrating normal and H-ras transformed 3T3 fibroblasts. Biophysical Journal. 80:1744-1757. (3) Engler, A.J., S. Sen, H.L. Sweeney, and D.E. Discher. 2006. Matrix elasticity directs stem cell lineage specification. Cell. 126:677-689. (4)Tan, J.L., J. Tien, D.M. Pirone, D.S. Gray, K. Bhadriraju, and C.S. Chen. 2003. Cells lying on a bed of microneedles: an approach to isolate mechanical force. Proc Natl Acad Sci USA. 100:1484-1489. |
【ひ】
| PAR-aPKC システム |
|---|
| 【PAR-aPKC system 】 |
| 鈴木 厚 |
| 横浜市立大学大学院医学研究科 分子細胞生物学 医学部分子生物学教室 |
| お問合せ |
| atypical PKC、および6種のPARタンパク質(PAR-1〜PAR-6)で構成される、種を超えて存在する普遍的細胞極性制御シグナル伝達システムを指す(線虫以外ではPAR-2に相当するタンパク質はいまだ同定されていない)。
細胞分化や幹細胞維持に必要な細胞の非対称分裂、さらには、分化を終えた上皮細胞や神経細胞の形態、機能の非対称化・極性化にも必須な機能を発揮している。多細胞生物の誕生とともに出現したシステムと考えられ、多細胞個体の複雑な形態形成と細胞特異的な機能獲得を可能にする上で、重要な役割をはたしてきたと推測される。 細胞内外からの極性シグナルに応答して、システム内のタンパク質間の相互作用にが生じ、細胞膜直下においてaPKC/PAR-3/PAR-6複合体とPAR-1が相互排除的に分布し、そのことによってまず、非対称な膜ドメインを確立される。その後に、それぞれの因子(特に、aPKC, およびPAR-1といったセリン・スレオニンキナーゼ)が細胞骨格系や細胞内輸送系を制御することによって細胞全体の非対称化が引き起されると考えられる。 参考文献A. Suzuki & S. Ohno The PAR-aPKC system: lesson in polarity. J. Cell Sci. 119: 979-987 (2006) 鈴木 厚、大野茂男, 細胞工学 24(3)、 「aPKC/PARシステム:多細胞生物の細胞極性を普遍的にコントロールする分子制御装置」227-230, 秀潤社, 2005 |
| ビンキュリン |
|---|
| 【vinculin】 |
| 木岡 紀幸 |
| 京都大学大学院農学研究科 |
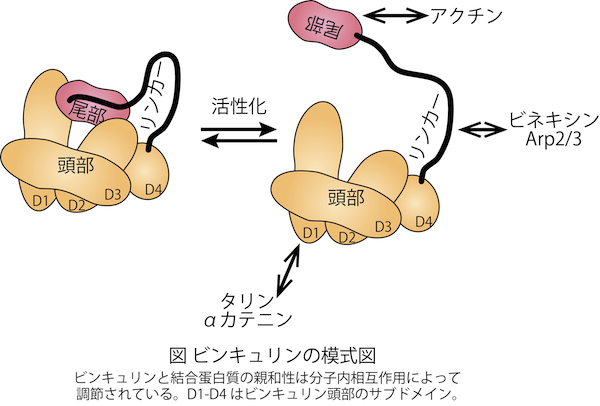
アクチン繊維と結合し、主に、細胞外マトリックス-細胞間の接着装置である接着斑(focal adhesion)と細胞-細胞間の接着装置であるアドヘレンス・ジャンクション(adherens junction)の両方に存在するタンパク質。ビンキュリンは頭部と尾部およびそれらをつなぐリンカー領域からなっており(図)、尾部にはアクチンが、頭部にはタリン(talin)やαカテニン(α-catenin)が結合する。タリンなどのタンパク質との相互作用を介して、ビンキュリンは接着斑の成熟やその接着強度に貢献している。リンカー領域にも細胞増殖シグナルに関わるビネキシン(vinexin)やインスリンのシグナル伝達に関与するCAP/ポンシン、アクチン重合の核形成因子Arp2/3が結合する。ビンキュリンとアクチンなどの結合タンパク質との親和性は頭部と尾部の分子内相互作用により低く抑えられており、分子内相互作用の解離に伴いその親和性が著しく増大する(活性化)。この活性化の仕組みは現在論争中であるが、複数の結合相手が同時にビンキュリンに作用することが活性化を引き起こすという「組合せモデル」が有力である。また接着斑でのビンキュリン-タリン結合およびアドヘレンス・ジャンクションでのビンキュリン-αカテニン結合が細胞内張力によって調節されることから、ビンキュリンはメカノトランスダクション(mechanotransduction)にも関与していると考えられている。参考文献Carisey, A. and Ballestrem (2011) Eur J Cell Biol 90 157-163. Kioka, N. (2002) Cell Struct Func 27 1-7. 木岡紀幸(2007) 生体の科学 58 430-431. |
| 非筋ミオシンIIのリン酸化による制御 |
|---|
| 【Regulation of non-muscle myosin II function by its phosphorylation】 |
| 米村 重信 |
| 理化学研究所 発生・再生科学総合研究センター (CDB) |
| お問合せ |
非筋細胞におけるミオシンIIはそのリン酸化によってATPase活性や会合能が制御されている。これは細胞の局所において、必要な時間だけミオシンIIが機能するためだと考えられている。常にアクチン繊維との相互作用によって収縮運動を行う骨格筋におけるミオシンIIではリン酸化による活性や会合能の制御が失われている。主要な調節を担うリン酸化を受けるのは生物種によって重鎖である場合と調節軽鎖(myosin regulatory light chain: MRLC)である場合があるが、ここではショウジョウバエから哺乳類まで広く見られるMRLCのリン酸化について述べる。哺乳類の場合、MRLCの19番目のセリン、18番目のスレオニンが順次リン酸化を受け、リン酸化を多く受ける程ATPase活性も会合したミオシンIIフィラメントも安定性も上昇することが生化学的にわかっている。ショウジョウバエではMRLCの変異体ではミオシンII活性に依存する細胞質分裂が異常になり、その異常はリン酸化されない変異MRLCでは回復せず、野生型あるいは疑似リン酸化型のMRLCの導入によって回復した。これらのことによりミオシンIIのリン酸化による制御の重要性がわかる。19番目のセリンのみがリン酸化されているMRLC (1P-MRLC)を認識する抗体、18番目のスレオニンと19番目のセリンと両方がリン酸化されているMRLC (2P-MRLC)を認識する抗体がそれぞれ作られている。野生型MRLC、リン酸化されない変異MRLC、擬似リン酸化型変異MRLCなどを細胞に発現させ、ミオシンIIのダイナミクスを観察すると、1P-MRLCは会合したミオシンIIと同じ局在を示すが、収縮しつつある場所には2P-MRLCが多いこと、ミオシンIIの細胞内の局所的な集積離散にはリン酸化脱リン酸化を繰り返すことのできるMRLCが重要であることがわかる。細胞内でリン酸化を行っているキナーゼにはミオシン軽鎖キナーゼ(MLCK)、ROCK/Rho-kinase、citron kinase、MRCK、ZIP kinaseなどが報告されており、阻害剤やノックダウン、ノックアウトの実験からその重要性が指摘されている。脱リン酸化の方はMYPT1というサブユニットを含むミオシンフォスファターゼが担当している。ROCK/Rho-kinaseは直接MRLCをリン酸化する能力があるとともに、MYPT1をリン酸化することによりミオシンフォスファターゼ活性を抑制し、細胞内のミオシンリン酸化に大きな役割を果たしていることがわかっている。参考文献 |
| 非筋ミオシンIIの会合 |
|---|
| 【Assembly of nonmuscle myosin II】 |
| 高橋 正行 |
| 北海道大学大学院理学研究院 |
| お問合せ |
ミオシンスーパーファミリーの中でミオシンIIの際立った特徴は、会合して双極性のフィラメント(bipolar filament)を形成することである。両端に頭部を向けた双極性のフィラメント構造をとることにより、極性をもつアクチンフィラメントを両方向からたぐり寄せられる。その結果、細胞内のアクチン細胞骨格を収縮させる、或いは骨格に張力をかけることができる。ミオシンIIの会合は分子同士の静電相互作用による。会合に必須な領域(Assembly Competence Domain: ACD)は重鎖(myosin heavy chain)のα-helical coiled-coil構造からなる尾部のC末端近傍にあることがわかってきている。非筋ミオシンII(nonmuscle myosin II)のフィラメントは横紋筋ミオシンの太いフィラメント(thick filament)よりもかなり小さく、15〜30個程の分子でできている。横紋筋ミオシンはサルコメア構造内で常に太いフィラメントを形成しているのに対し、非筋ミオシンIIは細胞内で然るべき時に然るべき部位に集まって会合し、アクチンフィラメントと共に収縮性の構造体を一時的に形成する。精製したミオシンIIのin vitroにおける解析から、ミオシンIIは不活性な状態では棒状の尾部が二箇所で折り畳まれた、会合できないコンフォメーションをとり、調節軽鎖(regulatory light chain)のSer19(場合によってはThr19も)がリン酸化されると、頭部のモーター活性が上昇するのと同時に尾部が伸びたコンフォメーションに変化し会合できるようになると考えられている。細胞性粘菌のミオシンIIのフィラメント形成は重鎖のリン酸化により調節されている。一箇所で折れ曲がった尾部が脱リン酸化により伸びて会合できるようになる。脊椎動物のミオシンIIも軽鎖のリン酸化に加えて、重鎖のリン酸化による調節機構もあることがわかってきている。尾部の重鎖C末端のACDの近傍とnonhelical tailpiece(α-helical coiled-coil構造がほどけた領域)と呼ばれる領域に、PKC、CK II、TRPM7等によるリン酸化部位が存在し、リン酸化により会合能が減少する。また、他のタンパク質との相互作用による会合調節機構も存在する。S100A4/Mts1はミオシンIIアイソフォーム(IIA、IIB、IIC)のうちIIAだけに特異的に結合し脱会合状態に保たせる。その結果、ガン化した細胞の転移能を増加させることが示唆されている。参考文献 |
| 非筋ミオシンIIの分子種の特徴 |
|---|
| 【Properties of nonmuscle myosin II isoforms】 |
| 高橋 正行 |
| 北海道大学大学院理学研究院 |
| お問合せ |
非筋ミオシンII(nonmuscle myosin II)は、アクチンフィラメントを動かす、或いはつなぎ止めることで、細胞の形態変化を含む様々な過程(細胞質分裂、細胞遊走、細胞間及び細胞−基質間接着等)に関与するモータータンパク質である。二本ずつの重鎖(myosin heavy chain:MHC)、必須軽鎖(essential light chain)、調節軽鎖(regulatory light chain)からなる六量体であり、二個の球状の頭部と長い棒状の尾部をもつユニークな構造をもつ。頭部は、 ATPase活性部位とアクチン結合部位を含むモータードメインと,軽鎖結合部位であるレバーアームからなり、アクチンフィラメントを動かすモータータンパク質としての機能を担う。尾部はミオシンIIが会合して双極性のフィラメントを形成することに関与する。ミオシンIIはフィラメントを形成して機能することにより、複数のアクチンフィラメントを同時につなぎ止める構造タンパク質としての役割も持つ。ほ乳類には三種類のミオシンII重鎖アイソフォーム(MHC-IIA,-IIB,-IIC)が存在し,それぞれがホモダイマーを形成してミオシンIIA,IIB,IICとなる。さらに、IIBとIICには頭部のLoop1(ATPase site 近傍)及びLoop2(アクチン結合部位近傍)に短い挿入配列をもつスプライスバリアント、B1、C1及びB2、C2が存在する。B1、B2、C2は神経組織特異的に発現している。IIAのノックアウトマウスは、近位内胚葉層形成の異常と細胞間接着の欠陥により胎生6.5日目で死亡する。IIBのノックアウトマウスは、その発現が多い組織である脳と心臓に発生異常が起こり、ほとんどが胎生14.5日目で死亡する。IIAは他のアイソフォームよりも高いモータータンパク質としての活性を示す。IIBは他のミオシンIIと比べて異常に高いduty ratio (1回のATP加水分解サイクルにおけるアクチンに結合している時間の割合)を示すことから、構造タンパク質としての役割が強い可能性もある。一方、IICは全ヒトゲノム塩基配列決定後、新たにその存在が明らかになったアイソフォームであるが、そのノックアウトマウスに顕著な異常は見られない。それぞれのアイソフォームにおいて、組織特異的な発現様式、また細胞内局在の違いも報告されており、重複した機能の他にアイソフォーム特異的な機能があることが考えられる。参考文献 |
| 非対称分裂 |
|---|
| 【Asymmetric cell division】 |
| 入江 賢児 |
| 筑波大学大学院人間総合科学研究科(基礎医学系)分子細胞生物学グループ |
| お問合せ |
細胞分裂によって2つの異なる細胞を生じるような細胞分裂。非対称分裂は細胞の多様性を生みだす基本的なしくみである。個体発生の過程では、少数の未分化な前駆細胞(もしくは幹細胞)が細胞増殖を繰り返し、特定の機能や構造を形成する細胞集団が形成されていく。非対称分裂は、このような過程で、細胞内の非対称性や細胞外の微細環境(ニッチ、niche)に基づいて、未分化な細胞からより分化した細胞を生じるメカニズムである。
参考文献1;Mechanisms of asymmetric cell division: two Bs or not two Bs, that is the question. Horvitz HR, Herskowitz I. Cell. 1992 Jan 24;68(2):237-55.
|
| 微小管のダイナミック・インスタビリティ |
|---|
| 【Dynamic instability of microtubules】 |
| 清末 優子 |
| 理化学研究所 神戸研究所 発生・再生科学総合研究センター (CDB) |
| お問合せ |
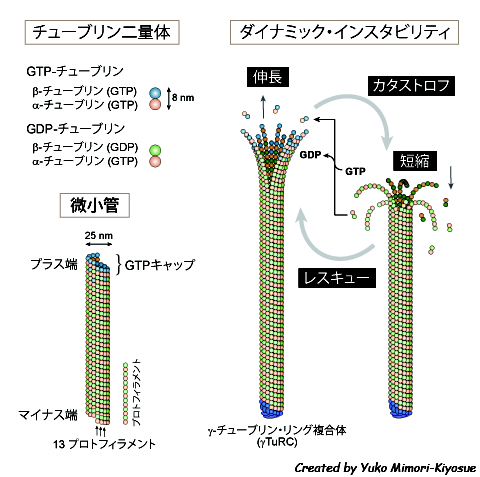
| 微小管は、α-チューブリンとβ-チューブリンのヘテロ二量体が重合して形成される、直径25 nmの筒状のフィラメントである(図)。α-/β-チューブリン二量体が同じ向きに長軸方向に共有結合してプロトフィラメントが形成され、13本のプロトフィラメントが側面で結合して微小管となる。微小管の伸長が早い端をプラス端、反対側をマイナス端とよび、プラス端先端にはβ-チューブリンが位置している。α-チューブリンとβ-チューブリンはそれぞれグアニンヌクレオチド結合部位を1個所もち、α-チューブリンは常にGTPを結合しているが、β-チューブリンのGTPは重合すると加水分解されてGDPとなり、脱重合してフリーになるとGDPが溶液中のGTPに交換されるというサイクルを繰り返している。GTPの加水分解はチューブリンの重合には必要ないが、GDP-チューブリンはフィラメントの外側に向けて反り返った構造をとり、脱重合しやすくなるため、フィラメントの速やかな崩壊をもたらす[1]。重合直後のまだGTPを保持しているチューブリンからなる先端部分は“GTPキャップ”とよばれ、脱重合を抑止して持続的な微小管伸長を助ける[2]。 個々の微小管の挙動を観察すると、精製タンパク質を用いたin vitro再構成系においても、細胞内においても、比較的ゆっくりとした伸長と、伸長よりも数倍早い速度での急速な短縮を繰り返している。この挙動はin vitroにおいても特にプラス端側で活発であるが、細胞内ではマイナス端側は安定化因子によってキャップされていることが多く非動的なため(「微小管マイナス端結合因子」の稿参照)、伸縮の繰り返しは主にプラス端側においてのみ観察される。フィラメントの端で自発的な重合と脱重合の相転換を起こすという性質は、細胞骨格ポリマーの中でも微小管に特徴的なもので、ダイナミック・インスタビリティ(動的不安定性)とよばれている[3, 4]。ダイナミック・インスタビリティのパラメータは、微小管の伸長(growth)と短縮(shrink)、伸長から短縮への変換をカタストロフ(catastrophe)、短縮から伸長への変換をレスキュー(rescue)で表す(図)。伸縮がない状態をポーズ(pause)とよぶが、チューブリンの重合・脱重合が全く生じないという状況はほとんど起こらないので、伸縮の繰り返しの距離が検出限界以下の状態ともいえる。また、系全体のダイナミクスさの程度を表す指標として、カタストロフとレスキューの頻度の総和としてダイナミシティ(dynamicity)という言葉で表すこともある。 このような動的な性質は、微小管ネットワークのパターンを必要に応じて速やかに再編するために役立つ[5]。細胞の中では、様々な因子がダイナミック・インスタビリティを調節し、微小管の長さや配置を時空間的に制御している(「古典的MAPs」「微小管不安定化因子」「微小管切断因子」「微小管の重合核形成」「微小管プラス端集積因子(+TIPs)」の稿参照)。微小管再編の最も顕著な例は、細胞分裂開始時、間期微小管が完全に崩壊して分裂期紡錘体が形成される劇的な過程であろう。間期においても、細胞構造に応じた多様な配置をとって、オルガネラの配置や物質輸送を担い、秩序だった細胞活動の基盤となっている。 参考文献1. Elie-Caille, C. et al. Straight GDP-tubulin protofilaments form in the presence of taxol. Current Biology 17, 1765-1770 (2007). 2. Howard, J. & Hyman, A.A. Growth, fluctuation and switching at microtubule plus ends. Nature Reviews Molecular Cell Biology 10, 569-574 (2009). 3. Hotani, H. & Horio, T. Dynamics of microtubules visualized by darkfield microscopy: treadmilling and dynamic instability. Cell Motil Cytoskeleton 10, 229-236 (1988). 4. Mitchison, T. & Kirschner, M. Dynamic instability of microtubule growth. Nature 312, 237-242 (1984). 5. Kirschner, M. & Mitchison, T. Beyond self-assembly: from microtubules to morphogenesis. Cell 45, 329-342 (1986). |
| 微小管の重合核形成 |
|---|
| 【Microtubule nucleation】 |
| 五島 剛太 |
| 名古屋大学大学院理学研究科生命理学専攻 |
| お問合せ |
| 微小管(microtubule)はチューブリン(tubulin)の重合体である。試験管内では高濃度のチューブリンとGTPを混ぜると重合反応が起きて微小管が容易に生み出されるが、細胞内では通常、チューブリンは高濃度では存在せず、偶発的な重合反応が起こることは稀である。重合の際に律速となっているのは最初の段階(=重合核の形成)である。いったんチューブリンの重合が開始されると(=重合核が形成されると)、チューブリンは低濃度でも効率よく次々と付加されて微小管は伸長する。微小管の重合核形成因子としては、γチューブリン複合体(γ-tubulin complex)が知られている。試験管内でγチューブリン複合体と低濃度のチューブリンを混ぜると微小管は効率よく生み出される。細胞内では、たとえばγチューブリン複合体が多く集積している中心体では、絶え間なく微小管が生み出されている。一方、細胞内に存在するγチューブリン複合体の大半を除去してもなお一部の微小管は作り出されることから、γチューブリン複合体とは別の重合核形成因子(nucleator)が存在することも示唆されている。 なお、筆者は「重合核形成」という言葉を用いているが、「nucleation」に対する日本語訳は定まっていない。実際の日常会話では日本語でもたいていそのまま「Nucleation(ヌクリエーション)」と言っている。 参考文献 |
| 微小管プラス端集積因子(+TIPs) |
|---|
| 【Microtubule plus-end-tracking proteins (+TIPs)】 |
| 清末 優子 |
| 理化学研究所 神戸研究所 発生・再生科学総合研究センター (CDB) |
| お問合せ |
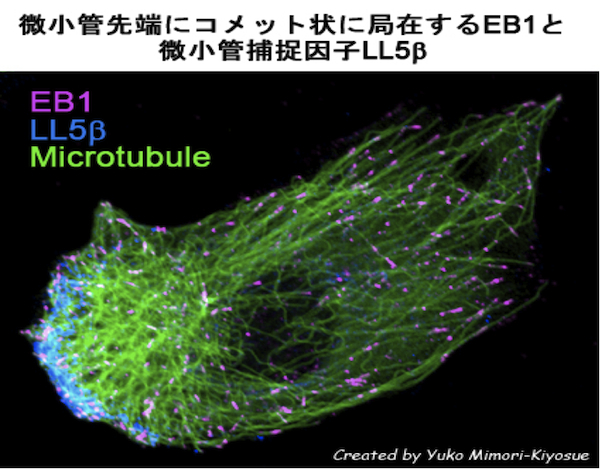
| 微小管マイナス端結合因子 |
|---|
| 【Microtubule minus-end binding proteins】 |
| 孟文 翔1・竹市 雅俊 2 |
| 1Institute of Genetics and Developmental Biology, Chinese Academy of Sciences, Beijing 2理化学研究所 神戸研究所 発生・再生科学総合研究センター (CDB) |
| お問合せ |
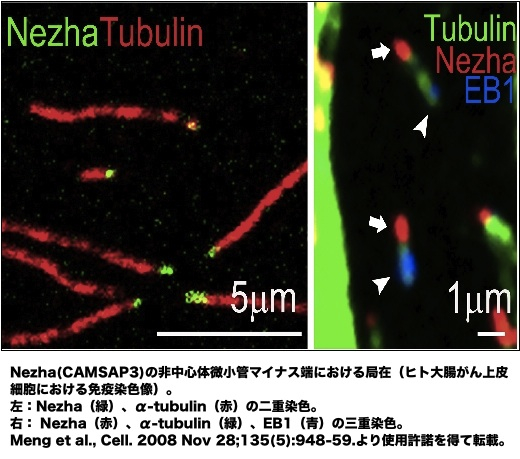
| 微小管は、プラス、マイナス両端において動的な構造体であるが、プラス端でチュ-ブリンの付加が起きやすい。この付加とマイナス端での解離がバランスされるとトレッドミル状態となることもあるが、マイナス端の脱重合速度はプラス端での重合速度よりも早いため、マイナス端が安定化されていなければ微小管は消滅する。しかし、マイナス端が何らかの形で安定化されると、微小管はブラス端だけで重合・脱重合を繰り返す[1, 2]。これが通常、細胞内でみられる微小管の状態である。 マイナス端の安定化は、中心体、またはそれ以外の細胞内構造(または分子)との結合によってもたらされ、それに対応して、微小管は中心体微小管、非中心体微小管とに分類される[2]。中心体では、γチューブリンがγ-チューブリン複合体蛋白質(γ-tubulin complex components, GCPs)と共にγチューブリン環複合体(γ-tubulin ring complex, γTuRC)を形成する。γTuRCは微小管マイナス端をキャップして安定化することで、プラス端における持続的な伸長/短縮ダイナミクスの繰り返しを可能とする。中心体は、さらに多数の成分を含み、微小管形成中心(microtubule organizing center, MTOC)としての役割を果たす[3-6]。哺乳類の細胞では、ninein、CEP192、CG-NAPとpericentrin などがγTuRCの機能を支え、これらの分子が欠損すると微小管の伸長が障害される[7-10]。 非中心体微小管は 上皮細胞、筋繊維、神経軸索等において顕著で、また、高等植物細胞はそもそも中心体を持たない。ゴルジ体、核膜、細胞膜等の構造体から非中心体微小管のプラス端重合が始まることが知られているが、その詳細な分子機構については未解明な部分が多い(繊毛微小管は、中心小体から成る基底小体から発するので、中心体微小管の一様式とみなすべきであろう)[2]。最近、微小管マイナス端結合因子Nezha(CAMSAP3)が同定され、非中心体微小管形成機構研究は新しい局面を迎えている。Nezhaは、接着結合(adherens junction, AJ)、及び、細胞質に分布し、微小管のマイナス端に結合してプラス端側の自由な伸長を支える(画像)。細胞接着部位では、Eカドヘリン-p120カテニン- PLEKHA7複合体にNezhaが結合することで微小管をAJにつなぎとめ、Nezha欠失によりAJが障害される[11, 12]。ショウジョウバエにはNezha 類似分子 Partronin(SSP4)があり、これが欠失すると細胞分裂時の紡錘体が縮小する。Partroninはマイナス端に結合することにより、キネシン13による微小管脱重合を阻害する[13, 14]。これは、Nezha/Patroninによる非中心体微小管の安定化機構をよく説明している。 またデスモソームにおいては、構成成分デスモプラキンに中心体から遊離したnineinが結合し、これが微小管をデスモソームにつなぎとめるという報告もあり、微小管マイナス端結合タンパク質が必要に応じて多様な細胞内構造に再配置される例が見つかってきている[15]。 参考文献1. Kollman, J.M., Merdes, A., Mourey, L., and Agard, D.A. Nature reviews Molecular cell biology 12:709-721 (2011)
2. Keating, T.J., and Borisy, G.G. Biol Cell 91:321-329 (1999) 3. Kollman, J.M., Polka, J.K., Zelter, A., Davis, T.N., and Agard, D.A. Nature 466:879-882 (2010) 4. Wiese, C., and Zheng, Y. Curr Opin Struct Biol 9:250-259 (1999) 5. Wiese, C., and Zheng, Y. Nat Cell Biol 2:358-364 (2000) 6. Murphy, S.M., Preble, A.M., Patel, U.K., O'Connell, K.L., Dias, D.P., Moritz, M., Agard, D., Stults, J.T., and Stearns, T. Mol Biol Cell 12:3340-3352 (2001) 7. Delgehyr, N., Sillibourne, J., and Bornens, M. J Cell Sci 118:1565-1575 (2005) 8. Gomez-Ferreria, M.A., Rath, U., Buster, D.W., Chanda, S.K., Caldwell, J.S., Rines, D.R., and Sharp, D.J. Curr Biol 17:1960-1966 (2007) 9. Takahashi, M., Yamagiwa, A., Nishimura, T., Mukai, H., and Ono, Y. Mol Biol Cell 13:3235-3245 (2002) 10. Zimmerman, W.C., Sillibourne, J., Rosa, J., and Doxsey, S.J. Mol Biol Cell 15:3642-3657 (2004) 11. Meng, W., Mushika, Y., Ichii, T., and Takeichi, M. Cell 135:948-959 (2008) 12. Akhmanova, A., and Yap, A.S. Cell 135:791-793 (2008) 13. Goodwin, S.S., and Vale, R.D. Cell 143:263-274 (2010) 14. Gardner, M.K., Zanic, M., Gell, C., Bormuth, V., and Howard. Cell 147:1092-1103 (2011) 15. Lechler, T., and Fuchs, E. J Cell Biol 176:147-154 (2007) |
| 微小管切断因子 |
|---|
| 【Microtubule Severing Factor】 |
| 岡田 康志 |
| 理化学研究所 神戸研究所 生命システム研究センター(QBiC) |
| 微小管切断因子(microtubule severing factor)とは、微小管を中途で切断する活性を持つ酵素である。椎名らの実験により細胞周期依存的な微小管切断活性の存在が示唆され、現在までに4種類の微小管切断酵素が同定されている。このうちEF1α以外の3種の蛋白は、膜輸送に関わるAAA蛋白VPS4と類縁で、katanin, spastin, fidgetin と名付けられている。このうちkataninは、分裂期細胞質中の微小管切断活性因子として同定され、日本語の刀に因んで命名された。Spastinおよびfidgetinは、それぞれ神経系の変異であるヒトの遺伝性痙性対麻痺(hereditary spastic paraplegia)およびマウスの自然変異体であるfidget(モジモジするという意味)の原因遺伝子として同定され、それぞれin vitroあるいはin vivoでの微小管切断活性が確認された。 変異体の解析により、これらの微小管切断因子は、細胞分裂における微小管の長さ・量の制御や、植物細胞の間期微小管の枝分かれ構造の形成・維持、線毛の調節、神経細胞での微小管の制御などの多様な機能を果たしていることが知られている。 VPS4自体には微小管切断活性は報告されていないが、リング状の12量体を形成して膜輸送蛋白複合体を解体する機能が知られている。リング状の6量体を形成するKataninやSpastinも、これと同様の機構で微小管を解体・切断すると提唱されている。 参考文献 |
| 微小管不安定化因子 |
|---|
| 【Microtubule Destabilizer】 |
| 岡田 康志 |
| 理化学研究所 神戸研究所 生命システム研究センター(QBiC) |
| 微小管(microtubule)は動的なポリマーで、その構成タンパクであるα/β チューブリン(tubulin)のヘテロ二量体の重合と解離により伸長(growth)と短縮(catastrophe)を繰り返す(動的非平衡性dynamic instability)。細胞内には、多数の伸長を促進する因子と短縮を促進する因子の双方が存在しており、組織や細胞あるいは細胞周期などに応じて微小管のダイナミクスが調節されている。このうち前者を微小管安定化因子(microtubule stabilizer)、後者を微小管不安定化因子(microtubule destabilizer)あるいは微小管崩壊因子(microtubule catastrophe factor)と総称している。
これまで3種類の異なるタイプの微小管不安定化因子が知られている。 1つは、微小管をその途中で切断する酵素で、微小管切断因子(microtubule severing factor)と呼ばれている。カタニン(katanin, 日本語の刀にちなんで名付けられた)がその代表である。 2つめのタイプは、微小管の端に局在し、端からの脱重合を促進する酵素で、微小管脱重合因子(microtubule depolymerizing factor)と呼ばれている。モータータンパク質(motor protein)のキネシン(kinesin)の一種であるkinesin-13 (M型キネシン、マウスのKIF2やヒトのMCAKなど)や、kinesin-8 (出芽酵母のKip3 など)が代表である。 3つめのタイプは、チューブリン二量体に高い親和性で結合することで微小管の重合脱重合の動的平衡を脱重合側へ移動する蛋白(チューブリン隔離因子, tubulin sequestering factor)で、スタスミン(stathmin, OP18)が代表である。ただし、結晶構造の比較などから、スタスミンもkinesin-13と同様の微小管端での脱重合促進活性を持つと考えられている。 また、微小管の重合を阻害する薬剤のコルヒチン(colchicine)やノコダゾール(nocodazole)なども、微小管不安定化因子と呼ばれることがある。 参考文献 |
【ふ】
| フィラミン |
|---|
| 【Filamin】 |
| 太田 安隆 |
| 北里大学理学部生物科学科 |
| お問合せ |
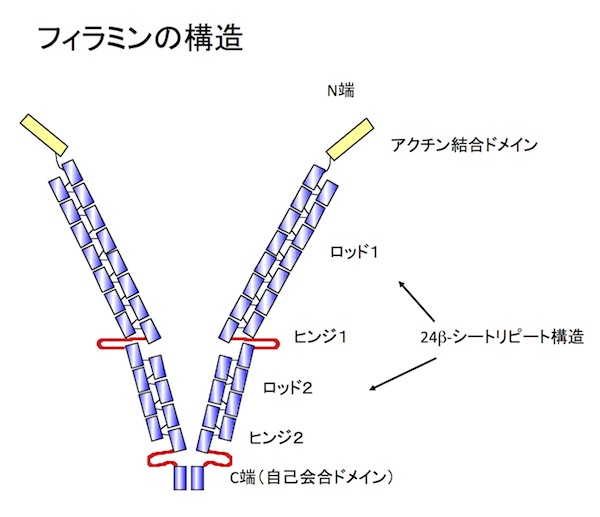
フィラミンは、アクチン繊維架橋タンパク質のひとつで、細胞皮層のアクチン繊維ネットワークの主要な構成分子の1つである。3種類のフィラミン(A, B, C)が知られている。このうちフィラミンAとBは各種臓器で発現してるが、フィラミンCは筋肉でのみ発現している。フィラミンは、分子量約280kDのサブユニットがC端で自己会合した2量体で、N端側のアクチン結合部位を使って、アクチン繊維を格子状に架橋してゲル構造を作ることができる(図1)。フィラミンAは、チャンネルタンパク質、受容体、タンパクキナーゼなど90種類以上の分子と結合し、シグナル伝達の足場タンパク質として機能している。血管内皮細胞の血流に対する応答や結合組織内を移動している白血球の運動など、細胞は様々な力学的な刺激に反応しながら生きている。フィラミンAは、細胞外からの力学的な力に応じてそのコンフォメーションを変化させ、インテグリンなどのフィラミン結合分子との相互作用を変化させることで力学的な刺激に対する細胞応答に関与することができる。フィラミン遺伝子の変異が複数の遺伝性疾患の原因であることがわかっている。例えばヒトにおいて、フィラミンの変異は脳室周囲異所性灰白室(フィラミンA)、家族制心臓弁膜ジストロフィー(フィラミンA)、boomerang dysplasia(骨異形成症)(フィラミンB)、筋原線維性ミオパチー(フィラミンC)を引き起こす。参考文献参考文献 Cell Adh Migr.5:160-9, 2011 (PMID: 21169733) |
| フィルギャップ |
|---|
| 【FilGAP】 |
| 太田 安隆 |
| 北里大学理学部生物科学科 |
| お問合せ |
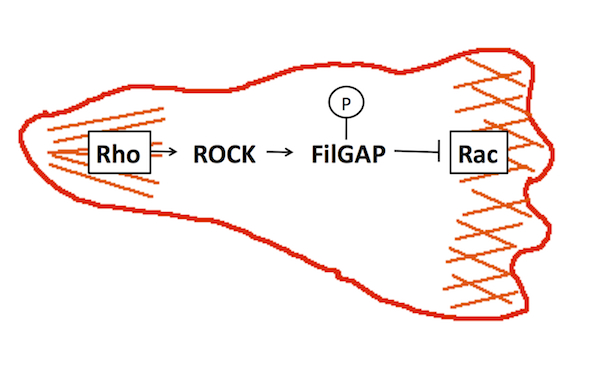
FilGAPは、低分子量GTP結合タンパク質 Racを特異的に不活化する因子GAP (GTPase-Activating Protein)の一つで、アクチン繊維架橋因子フィラミンに結合する。FilGAPを欠失した細胞は、細胞周囲に葉状仮足をランダムに進展させることからFilGPAは葉状仮足の形成制御に関与していると考えられる。FilGAPは、低分子量GTP結合タンパク質Rhoの標的タンパク質であるROCKキナーゼでリン酸化されると活性化される。細胞運動においては、Racが細胞の前部で活性化され葉状仮足を進展し、Rhoはミオシンを活性化し、収縮を促すことで細胞尾部を決定している。この細胞前部と尾部でのRacとRhoの特異的な活性化は、両者が互いに阻害し合っているからだと考えられており、FilGAPはRhoの下流でRacを不活化することで、RacとRhoの棲み分けに関与していると考えられる(図1)。FilGAPは、構造の似た他のRacGAPとファミリーを形成しており,メンバーとしてARHGAP22とARHGAP25が知られている。FilGAPの研究は北里大学理学部生体機能学講座のホームページに詳しく紹介されている。参考文献参考文献 Nat Cell Biol. 8:803-814, 2006 (PMID: 16862148) |
| フェルスター共鳴エネルギー移動 あるいは 蛍光共鳴エネルギー移動 |
|---|
| 【FRET (Förster Resonance Energy Transfer あるいはFluorescence Resonance Energy Transfer) 】 |
| 松田 道行 |
| 京都大学大学院生命科学研究科 生体制御学分野 |
| お問合せ |
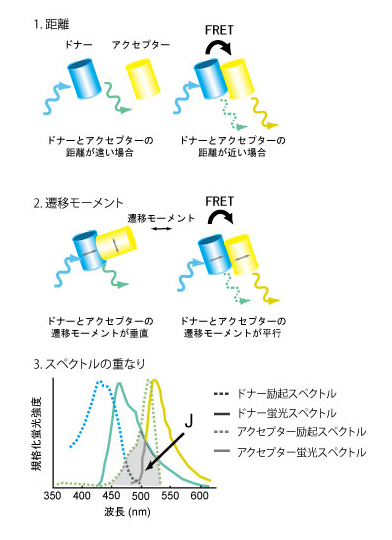
FRETとはこの現象の発見者の名前にちなんだFörster Resonance Energy Transferの略である。しかし、最近ではFluorescence Resonance Energy Transferの略と書いてある書籍が多く、蛍光共鳴エネルギー移動と邦訳されることが多い。FRETは、光子により励起されたドナー蛍光分子からごく近傍にあるアクセプター分子へエネルギーが共鳴移動する現象のことである。FRETの効率は、【1】蛍光発色団間の距離、【2】遷移モーメントの向き、【3】ドナー分子の蛍光波長域とアクセプター分子の吸光波長域の重なり、により主に規定される(1)。もっとも一般的なFRETの観察法はドナー蛍光の減少とアクセプター蛍光の増加を測定するが、厳密には、FRETはドナー分子の蛍光寿命を測定して定量される。FRETが蛍光タンパク質間でも観察されることを利用して、カルシウムを始めとするイオン、タンパク質リン酸化酵素、低分子量GTP結合タンパク質など、非常に多くの細胞内情報伝達分子に対するバイオセンサーが開発されている(2)。創薬においては、非常に長い蛍光寿命を持つユーロピウム(Eu)をドナーに用いることにより高いシグナルノイズ比のアッセー系が開発されている。参考文献 |
| フォルミンファミリータンパク質 |
|---|
| 【Formin homology proteins】 |
| 渡邊 直樹 |
| 東北大学 大学院生命科学研究科 |
| お問合せ |
フォルミンファミリータンパク質(以下、フォルミン)は、真核生物に広く存在するアクチン重合核形成促進因子である。哺乳類では〜15個、シロイヌナズナでは20余りの遺伝子が存在する。フォルミンの多くは、細胞質分裂、細胞極性形成に必須な遺伝子の産物である。マウスでは、Formin2の欠損で卵の減数分裂不全が、mDia1/3のダブルノックアウトで脳室下帯からの神経細胞の遊走の障害と、一部の個体において神経上皮の極性異常を伴った脳室周囲の異形成が生ずる。フォルミンは、C 末端側にフォルミン相同ドメイン1 と2(以下FH1,FH2)をもつ。FH1は、ポリプロリン配列の繰り返しからなり、FH2はフォルミンに特有のドメインで約400アミノ酸からなる。FH2はアクチン重合核形成作用をもつが、重合核形成後もアクチンの速い伸長端である反矢じり端(barbed end)にとどまり、連続的にアクチンを伸長させながら移動する、プロセッシブアクチン重合の性質をもつ。FH1は、複数のプロフィリン-アクチン複合体に結合することで、アクチン伸長を数倍に加速する。細胞内では、毎秒720個のアクチン単量体を取り込む速度で線維伸長することが、mDia1の細胞内分子可視化によって観察されている。参考文献 |
| プロフィリン |
|---|
| 【Profilin】 |
| 渡邊 直樹 |
| 東北大学 大学院生命科学研究科 |
| お問合せ |
12-15 kDのアクチンモノマー結合タンパク質。細胞質に豊富にある。単量体アクチンのADP-ATP交換反応を加速するとともに、ATP結合単量体アクチンに強く結合する(解離定数 〜0.1 μM)。プロフィリンに結合したアクチンは、速い重合端であるアクチン線維の反矢じり端(barbed end)に単量体アクチンと同様の速さで付加されるため、プロフィリンはアクチン伸長を助けるが、アクチン重合核の形成に対しては阻害的に働く。また、平衡状態のアクチン線維では脱重合端となる矢じり端(pointed end)への重合は阻害する。プロフィリン-アクチン複合体は、細胞質に10〜40 μM存在し、迅速な線維伸長を可能とする。また、アクチン伸長端に結合するフォルミンファミリータンパク質やEna/VASPタンパク質のポリプロリン配列に結合することで、協調して線維伸長を数倍加速する。インビトロでは、ADPアクチンの線維の反矢じり端からの脱重合を促進することも知られている。ホスファチジルイノシトール(4, 5)二リン酸などのホスホリピッドに結合するとアクチンとの相互作用が弱まる。参考文献 |
| 分子モーター |
|---|
| 【Molecular Motor】 |
| 岡田 康志 |
| 理化学研究所 神戸研究所 生命システム研究センター(QBiC) |
| 分子モーターとは、動きを作り出す生体分子の総称である。多くの場合、化学的エネルギーを力学的仕事に変換することで動きが作り出される。分子モーターとして特によく研究されているのが、細胞骨格系の上を動く分子モーターである。アクチン線維(F-actin)の上を動くミオシン(myosin)がその代表で、筋肉の運動をはじめ様々な生体運動に関わっている。また、微小管(microtubule)の上を動く軸糸ダイニン(axonemal dynein)は、真核細胞の線毛・鞭毛の運動を担うモーターである。キネシン(kinesin)や細胞質ダイニン(cytoplasmic dynein)も微小管上を動く分子モーターで、細胞分裂や細胞内の物質輸送、細胞内小器官の輸送や位置固定などを行っている。これらはいずれも、アデノシン三リン酸(ATP)をアデノシン二リン酸(ADP)に加水分解する酵素で、この反応によって得られる化学エネルギーを力学的仕事に変換することで運動を作り出す。
一方、バクテリアの鞭毛は細胞膜内外の電気化学ポテンシャル(H+やNa+の濃度勾配)を利用して回転運動を作り出すモーターにより駆動されている。ATP合成酵素のFoサブユニットも同様の機構で回転すると考えられている。ATP合成酵素のF1サブユニットは、Foサブユニットの回転を利用してATPを合成する酵素だが、その逆反応としてATPをADPに加水分解することでF1サブユニット自体が回転運動を作り出すことができる。 さらに、細胞骨格自体も、化学反応と共役した伸長・短縮を繰り返すことで力学的仕事を行っている。たとえば、細胞膜直下でのアクチン線維の重合は細胞膜伸展の駆動力であると考えられている。また、染色体の微小管の短縮に共役した輸送も知られている。 更に最近では、DNAやRNAの重合や修飾を行う酵素も分子モーターとしての活性を持つことが示されている。たとえば、DNAポリメラーゼ(polymerase)やRNAポリメラーゼ、あるいはリボソームは、DNAやRNAの重合、蛋白の翻訳に伴って鋳型DNA/RNAの上を移動する。ヘリケース(helicase)はDNA二本鎖を解離しながら移動し、ウイルスにはDNAをウイルスのカプシド(capsid)内に押し込むためのpackaging enzymeが存在する。さらに、クロマチン・リモデリング(chromatin remodeling)や、染色体形成(chromosome condensation)も力学的仕事を伴う過程だが、やはりATPの加水分解をエネルギー源とするモーター分子(SWI/SWF, SMCなど)が関与している。 ユニークな分子モーターとしては、ピエゾ素子のように電位依存的に内耳外有毛細胞を高速に伸縮させることで聴覚の感度調節を司るprestinがある。 参考文献 |
【へ】
| Heparin-binding EGF-like growth factor (HB-EGF) |
|---|
| 【Heparin-binding EGF-like growth factor (HB-EGF) 】 |
| 目加田 英輔 |
| 大阪大学微生物病研究所細胞機能分野 |
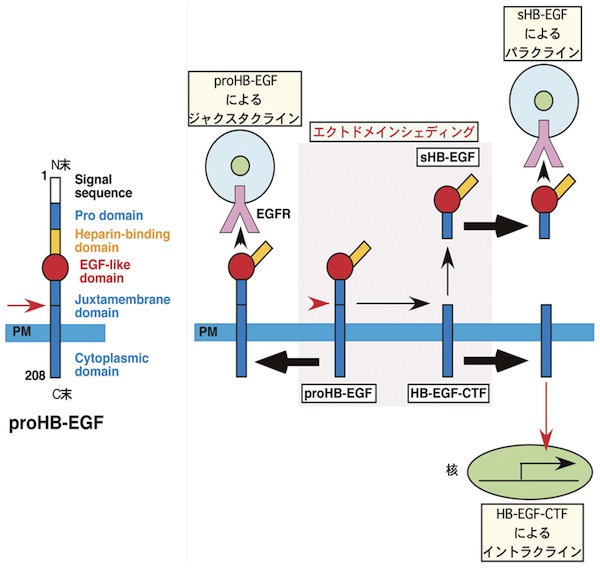
HB-EGFはEGFファミリーに属するヘパリン結合性の増殖因子である。この増殖因子は他のEGFファミリーと同様に、はじめ膜型蛋白質(proHB-EGF)として合成される。ProHB-EGFは細胞接着を介した細胞間情報伝達(ジャクスタクライン)に機能する一方、ジフテリア毒素受容体としても機能する。ProHB-EGFは細胞表面において種々の刺激に伴って酵素的切断を受け(エクトドメイン・シェディング)、その分泌型(sHB-EGF)が細胞外に放出される。sHB-EGFはEGF受容体(EGFR/ErbB1)あるいはErbB4に結合し、細胞増殖・運動など種々のシグナルを伝達する。シェディングに伴ってsHB-EGFとともに生成されるC末断片(HB-EGF-CTF)も細胞内へ移行して種々の遺伝子発現に関与する。HB-EGFノックアウトマウスなど種々の変異マウスの解析から、HB-EGFは心臓の発生や機能維持など様々な生理的過程、あるいは発癌などの様々な病理的過程で重要な働きをしていることが明らかとなっている。
参考文献UCSD-Nature Molecule Pages doi: 10.1038/mp.a002932.01, 2008 蛋白質核酸酵素 54, 1722-1727, 2009 |
【ほ】
| 紡錘体形成 |
|---|
| 【Spindle assembly】 |
| 五島 剛太 |
| 名古屋大学大学院理学研究科生命理学専攻 |
| お問合せ |
紡錘体(スピンドル:spindle)は細胞分裂の際に形成される双極性(bipolar)の構造体であり、細胞分裂装置(mitotic apparatus)と呼ぶこともある。染色体を娘細胞へと分配し、細胞質分裂を制御するために必須の役割を果たす。主要構成因子はチューブリン(tubulin)の重合体である微小管(microtubule)、および、さまざまな微小管結合蛋白質(microtubule-associated protein: MAP)である。動物体細胞においては2つの中心体(centrosome)が主な微小管形成中心(microtubule organising centre: MTOC)となり、中心体から伸びた微小管が凝縮した染色体を捕捉する。一方、植物細胞や減数分裂期の動物細胞の一部には中心体が存在せず、別のメカニズムにより紡錘体微小管が生み出されている。これまでに、染色体自身が微小管の形成を誘導することや(Ran GTPアーゼやAurora Bキナーゼが関わる)、既存の微小管に依存して新たに微小管が生み出される微小管増幅経路(オーグミン複合体が関与する)の存在が明らかにされた。また、動物体細胞においてもこうした経路が中心体経路と同時に働いていることも明らかになった。参考文献 |





